
ОПИСАНИЕ:
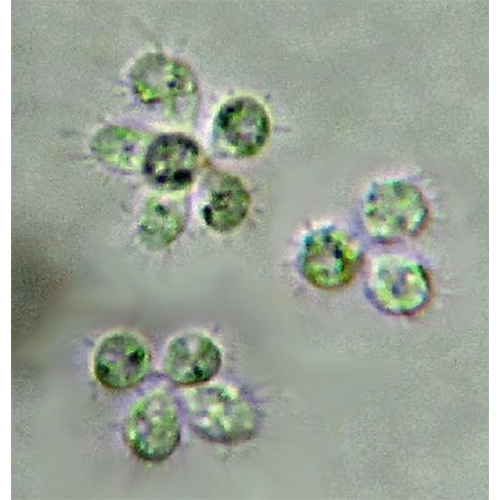
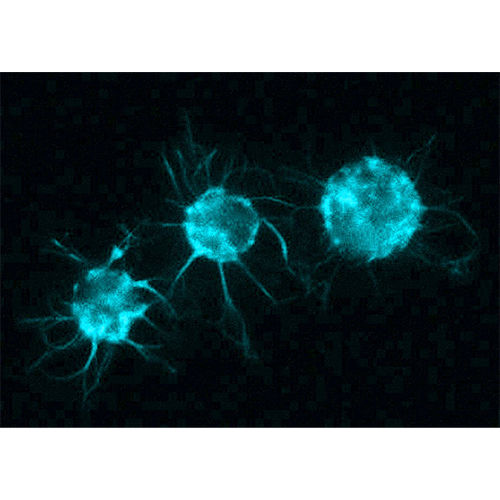
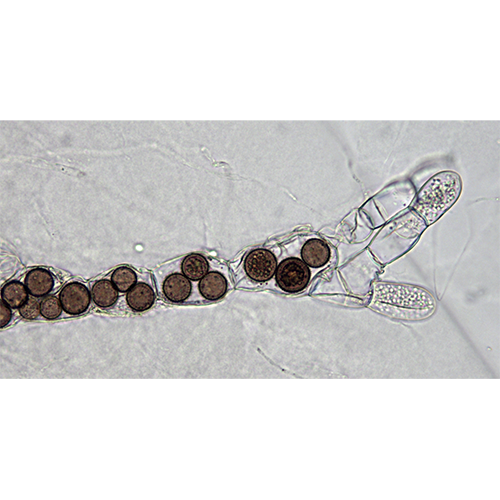
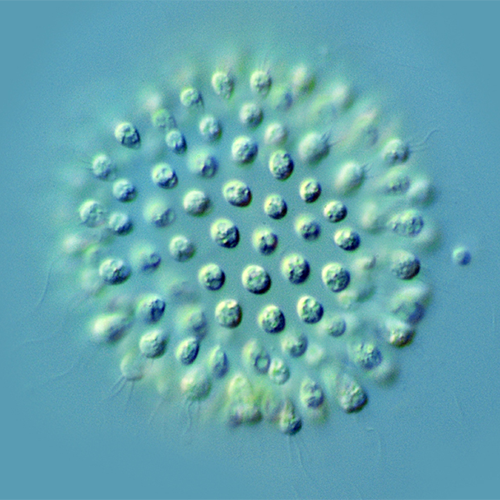
Хоанофлагеллаты, или воротничковые жгутиконосцы (лат. Choanoflagellata), — тип одноклеточных эукариот (протистов). Характерная черта — наличие у клетки одного жгутика, окружённого воротничком из 30—40 микроворсинок. Хоанофлагеллаты представлены одиночными и колониальными формами, обитающими в морских и пресных водах. Описано около 150 видов.
Научное название таксона образовано от др.-греч. χοάνη — воронка (по форме воротничка) и лат. flagellum — жгутик.
Строение
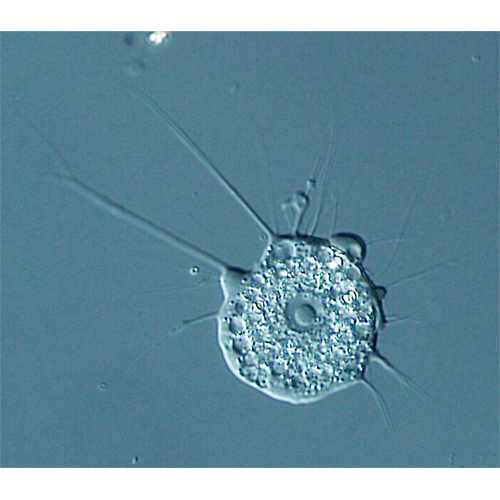
Размер хоанофлагеллат обычно превышает 10 микрон. Клетки демонстрируют отчётливую полярность, которая задаётся положением единственного жгутика. У многих представителей противоположным концом клетка прикрепляется к субстрату или погружена во внеклеточный матрикс колонии. Некоторые хоанофлагеллаты способны выделять бокаловидные «домики», которые у морских форм состоят из изящно переплетённых между собой кремниевых нитей. Пресноводные представители строят домик из целлюлозы.Жгутик и воротничок
Жгутик окружён венчиком (воротничком) из плотно примыкающих друг к другу микроворсинок (микровиллей) — пальцевидных выростов клетки, укреплённых актиновым цитоскелетом. Биение жгутика создаёт токи жидкости, направленные вдоль него от клетки. При этом вода постоянно поступает внутрь воротничка, проходя между микровиллей. Эти токи используются хоанофлагеллатами при питании: крупные частицы (бактерии, детрит) отцеживаются микровиллями и могут быть поглощены путём фагоцитоза.У плавающих в толще воды хоанофлагеллат движение жгутика также используется для локомоции. При этом организм перемещается жгутиком назад, подобно другим представителям группы Opisthokonta — грибам и многоклеточным животным.
Расшифровка генома
В 2008 году был полностью расшифрован геном одного из видов хоанофлагеллат — Monosiga brevicollis. В геноме моносиги около 9200 генов, причем присутствуют 23 гена кадгеринов и ряд других генов, типичных для многоклеточных животных, но отсутствующих у большинства других групп протистов.Филогения
Cходство воротничкового аппарата хоанофлагеллят с аналогичными структурами хоаноцитов губок может быть объяснено только их общим происхождением. На основании этого в прошлом некоторые авторы предполагали, что губки (Porifera) произошли от одноклеточных воротничковых жгутиконосцев, а прочие многоклеточные животные произошли от каких-то иных одноклеточных жгутиконосцев, не имеющих воротничка. Таким образом, было высказано предположение о полифилии многоклеточных животных – Metazoa. Однако в настоящее время известно, что у некоторых Monostomata (= Eumetazoa) имеются клетки, сходные по строению с хоаноцитами – циртоциты; они найдены в отдельных тканях на определенных стадиях развития у некоторых кишечнополостных (Cnidaria), немертин (Rhynchocoela), кольчатых червей (Annelida), моллюсков (Malacozoa), иглокожих (Echinodermata) и некоторых других; у многих Monostomata обнаружены клетки, произошедшие от воротничковых и сохраняющие элементы воротничка, но модифицированные для выполнение иных функций – сенсорной, прикрепительной или иной (Norrevang & Wingstrand 1970, Cantell et al. 1982). На основании этого делается вывод, что все Metazoa исходно имели хоаноциты и произошли от воротничковых жгутиконосцев. У воротничковых жгутиконосцев и у губок воротничок имеет консервативное строение; вероятно, у Monostomata произошла деконсервация строения воротничка, в результате чего он подвергся различным эволюционным преобразованиям и во многих случаях вообще исчез. В соответствии с этим предполагается родство между Metazoa и Choanoflagellata и высказано предложение объединить Metazoa и Choanoflagellata в таксон Apoikozoa Budd & Jensen, 2017 или объединить их с некоторыми другими одноклеточными в таксон Filozoa Shalchian-Tabrizi et al. 2008. Некоторые авторы вместо Apoikozoa неверно употребляют название «Choanozoa» (что не соответствует первоначальному значению названия Choanozoa Cavalier-Smith 1987, которое является синонимом названия Choanoflagellata). Границы таксонов Apoikozoa и Filozoa, которые характеризуются исходным наличием воротничка или образующих его отростков, неясны, поскольку не исключено, что у некоторых одноклеточных воротничок вторично исчез, так же, как он исчез у большинства Monostomata.Одиночные или колониальные гетеротрофные жгутиконосцы, планктонные или прикрепленные. Клетки овальной или грушевидной формы, размером от 3 до 25 мкм. На апикальном конце клетки находится 1 жгутик, окруженный тентакулами (микровиллями), образующими воротничок. В переходной зоне жгутика имеется центральный филамент, микротрубочковые корешки расходятся радиально от жгутиковой кинетосомы, кристы в митохондриях пластинчатого типа. Особи могут жить в домиках или без них, часто обладают хорошо развитым гликокаликсом. Ядро 1, расположено в центре или в передней половине клетки.
Размножаются, по-видимому, только бесполым путем преимущественно за счет продольного деления клетки на 2 дочерние. В жизненном цикле обычно есть 2 основные формы: прикрепленная или свободноплавающая и стадия покоя — циста. Распространены во всех районах земного шара; встречаются в пресных, солоноватых и морских водах. Свободноживущие.
Эта группа — одна из ведущих среди зоофлагеллат в планктоне, бентосе и обрастаниях. Их численность и биомасса в определенные сезоны соизмеримы с таковыми бактерий и зоопланктона. Обладая большой скоростью размножения и являясь активными бактериофагами, воротничкЪвые жгутиконосцы играют значительную роль в жизни водоемов.
История изучения
Впервые воротничковые жгутиконосцы (или хоа- нофлагеллаты) были описаны Эренбергом (Ehrenberg, 1838), ошибочно принявшим их за инфузорий из рода Epistylis. Этот вид впоследствии был назван Со- donosiga botrytis Stein, 1878. В 1868 г. Джеймс-Кларк (James-Clark, 1868) описал еще несколько видов хоа- нофлагеллат и впервые выделил их в особую группу Craspedomonadines. Однако статус таксономической категории этим простейшим — «Отряд Choanoflagel- lata» — впервые придал Кент (Kent, 1880—1882). С тех пор воротничковые жгутиконосцы изучаются как единая, четко очерченная группа и альгологами, и зоологами. Дальнейшие исследования хоанофлагеллат можно разделить на 2 периода: светооптический (до 1954 г.) и электронно-микроскопический.Использование только световой оптики не позволяло выяснить многие морфологические особенности этих мелких жгутиконосцев, в частности строение воротничка. В первый период было описано много новых видов и родов и начаты исследования питания и размножения.
Применение электронного микроскопа дало возможность установить строение воротничка, уточнить способы захвата и заглатывания пищи, углубить представления о способах размножения хоанофлагеллат. В результате изучения органелл клетки были сделаны важные обобщения, касающиеся не только этой группы, но и жгутиконосцев в целом.
Первая работа на современном уровне была сделана Хиббердом на Codonosiga botrytis (Hibberd, 1975). Серию исследований по ультраструктуре и жизненному циклу провели Ледбите с соавт. (Leadbeater, Morton, 1974; Leadbeater, 1977, 1983, 1991, 1994) и Карпов с соавт. (Карпов, 1981, 1982а, 19826, 1985; Жуков, Карпов, 1985; Карпов, Мыльников, 1993; Карпов, Ледбите, 1997, 1998). Видовое многообразие хоанофлагеллат, особенно, морских видов, исследовали Трондзен (Throndsen, Г974), Томсен с соавт. (Thomsen, Buck, 1991, Thomsen et al., 1995, 1997) и Мар- шан (Marchant, 1990).
Эти и другие работы подтвердили морфологическое единообразие в строении клетки хоанофлагеллат, а также своеобразие и четкую очерченность этого таксона. В настоящее время совершенно неясны связи этой группы с другими жгутиконосцами. Поэтому в дальнейшем необходимо расширить ультраструктур- ные исследования морских видов (акантоецид).
Спорным остается вопрос о родстве хоанофлагеллат и многоклеточных животных, особенно губок. Однако здесь не хватает данных по строению жгутиковых клеток метазоев.
Классификация
Четкая очерченность таксона и своеобразие в строении хоанофлагеллат побуждают исследователей повышать ранг этого таксона до типа (Corliss, 1984), надтипа (Карпов, 1990) и даже подцарства (Mohn, 1984). В настоящем руководстве принимается ранг типа Choanomonada, который представлен 1 классом Choanomonadea Kent, 1880, содержащим единственный отряд Choanoflagellida Kent, 1880. Отряд делится на 3 семейства, представители которых различаются по наличию или отсутствию домиков и их строению.Сем. Monosigidae Kent, 1880 — клетки без домиков, часто погружены в слизь.
Сем. Salpingoecidae Kent, 1880 — клетки в домиках из целлюлозы.
Сем. Acanthoecidae Norris, 1965 — клетки в домиках из кремниевых волокон.
Морфология
Клетки воротничковых жгутиконосцев независимо от принадлежности к семейству, а также у одиночных и колониальных видов устроены сходно. Тело особи имеет грушевидную или овальную форму. Сужение его передней части называется шейкой. Из апикальной части шейки выходит жгутик, окруженный воротничком. Многие хоанофлагеллаты имеют домики, которые бывают 2 типов: прозрачные из целлюлозы и в форме корзинки из жестких кремниевых полосок, или волокон (лорика). Особи без домиков обычно окружены слизью или тонкой оболочкой, которую можно обнаружить только под электронным микроскопом.Оболочка сохраняется у текатных и лорикатных хоанофлагеллат для фиксации клетки в домике.
Воротничковые жгутиконосцы снабжены наружными выростами, к которым относятся жгутик, окружающие его тентакулы воротничка, тентакулообразные выросты в других местах клетки и цитоплазматические мостики между соседними особями в колониях некоторых видов.
Тентакулы воротничка представляют собой тонкие цитоплазматические выросты толщиной около 200 нм. Они примерно одинаковой длины и разделены равными промежутками, располагаясь вокруг жгутика по окружности. Внутри каждой тентакулы проходит пучок актиновых микрофиламентов, поддерживающий ее в расправленном состоянии.
Тентакулообразные выросты встречаются и в других частях клетки. Чаще они располагаются на заднем конце особи и служат для прикрепления к субстрату или образования стебелька. Иногда есть и боковые выросты. У текатных и лорикатных форм они принимают участие в образовании домика. Тентакулообразные выросты могут быть временными или постоянными, а число их значительно меняется в пределах онтогенеза одной клетки. Внутри них также есть актиновые микрофиламенты, что свидетельствует о гомологии всех тентакулообразных выростов Хоанофлагеллат. Тентакулы воротничковых сходны с микрофиллями некоторых специализированных клеток многоклеточных животных. И те и другие имеют постоянную толщину по всей длине, расположены примерно на одинаковом расстоянии друг от друга, равны по длине и содержат микрофиламенты, состоящие из актина.
Латеральные цитоплазматические мостики между соседними клетками обнаружены в колониях Codonosiga botrytis, Desmarella moniliformis и D. thienemanni. Это временные образования, остающиеся, вероятно, после не прошедшего до конца деления клеток. Внутри мостика обычно имеется плотная перегородка, которая пронизана 1 или несколькими порами. Вероятно, цитоплазмы соседних клеток колонии еще некоторое время сообщаются между собой через поры.
Строение свободной части жгутика типично для большинства подобных образований. Это длинный цитоплазматический вырост апикального конца клетки, внутри которого проходит аксонема, состоящая из 2 центральных микротрубочек и 9 периферических дублетов. У некоторых видов на жгутике обнаружены очень тонкие боковые волоски — простые мастигонемы. Однако у большинства изученных воротничковых жгутиконосцев мастигонемы отсутствуют. Строение переходной зоны хоанофлагеллат уникально. Центральные микротрубочки расположены значительно выше поперечной пластинки и связаны с ней центральным филаментом.
Базальное тело жгутика, или кинетосома, состоит из 9 триплетов периферических микротрубочек. Дистальная часть кинетосомы соединяется с поверхностной мембраной в основании жгутика при помощи хорошо выраженных 9 переходных фибрилл. Так устроена жгутиковая кинетосома. Ниже и перпендикулярно к ней расположена безжгутиковая кинетосома, не отличающаяся по строению от обычной центриоли. Обе кинетосомы связаны друг с другом фибриллярным мостиком. Базальный комплекс жгутика, по-видимому, у всех хоанофлагеллат имеет диплосомное строение, т. е. состоит из 2 ортогонально расположенных кинетосом.
Корешковая система жгутиков воротничковых жгутиконосцев представлена лентами микротрубочек, которые радиально .расходятся от жгутиковой кинетосомы и лежат в одной плоскости с ее продольной осью. Ленты микротрубочек начинаются от сателлитов — электронноплотных образований, расположенных вокруг кинетосомы. Число и форма сателлитов у разных видов могут быть различны. Различается и количество микротрубочек. Например, почти у всех одиночных видов общее число микротрубочек в корешковой системе не превышает 100, а у колониальных оно может достигать 200—250. Микротрубочки расходятся под поверхностной мембраной, образуя скелет передней половины клетки и одновременно защищая снаружи ядро. Проходя также между основаниями тентакул воротничка, они обычно контактируют с пучками их микрофиламентов и формируют, таким образом, единый цитоскелетный комплекс. От безжгутиковой кинетосомы отходит тонкий поперечно исчерченный фибриллярный корешок, с которым связана мембрана сильно уплощенного мешочка. Весь этот фибриллярно-мембранный комплекс направлен к диктиосоме и обычно с ней связан.
Ядро воротничковых жгутиконосцев относится к пузырьковидному типу. Оно лежит в передней половине клетки. Ядрышко довольно крупное, занимает центральное положение. Глыбки гетерохроматина немногочисленны, распределены по всему объему ядра.
Между ядром и базальным комплексом жгутика всегда расположена единственная диктиосома аппарата Гольджи . Митохондрии с пластинчатыми кристами чаще находятся вблизи ядра. 1 или 2 сократительные вакуоли лежат в задней части клетки недалеко от поверхности. У морских видов они могут отсутствовать.
В задней же половине скапливаются пищеварительные вакуоли и включения запасных питательных веществ (липидов). У одиночных видов пищеварительные вакуоли обычно мелкие. В них содержится по 1—3 бактерии. Клетки колониальных видов часто имеют крупные пищеварительные вакуоли (могут содержать до 20 бактерий). Эти различия связаны с различиями в способах питания хоанофлагеллат (см. ниже). Цитоплазма довольно плотная, с большим количеством рибосом. У морского вида Choanoeca регplexa были обнаружены немногочисленные симбиотические бактерии (Leadbeater, 1977).
Подводя итог морфологическому описанию воротничковых жгутиконосцев, следует отметить, что все они устроены весьма однообразно. Сначала это заключение было основано на светооптических наблюдениях, а затем полностью подтвердилось электронно-микроскопическими исследованиями. Это особенно важно потому, что выделяемая ранее только по наличию воротничка группа простейших оказалась однородной и по многим другим признакам: у всех хоанофлагеллат между ядром и базальным комплексом жгутика лежит аппарат Гольджи; митохондрии с пластинчатыми кристами; жгутик 1, кнутовидного типа; базальный аппарат имеет диплосомное строение; в переходной зоне жгутика расположен центральный филамент; корешковая система жгутика образована радиально расходящимися лентами микротрубочек и 1 фибриллярным корешком. Практически все воротничковые жгутиконосцы способны образовывать временные или постоянные выросты, морфологически сходные с тентакулами воротничка. Поразительное однообразие ультраструктурной организации хоанофлагеллат, вероятно, свидетельствует об их филогенетическом единстве.
Питание
Воротничок хоанофлагеллат всегда и справедливо считался органоидом питания. Однако объясняли этот процесс по-разному. Одним из примеров может служить работа Кента (Kent, 1880—1882). Автор полагал, что пищевые частицы попадают внутрь воротничка и заглатываются у основания жгутика. В дальнейшем эти предположения не подтвердились. Верным оказалось лишь то, что жгутик создает направленные токи жидкости вокруг неподвижной клетки. Движение жидкости осуществляется таким образом, что увлекаемые им пищевые частицы попадают на наружную поверхность воротничка и прилипают к ней. В районе шейки простейшего появляется язычковидная псевдоподия, которая растет по направлению к воротничку. Она захватывает пищевые частицы с наружной поверхности воротничка и, замыкаясь, образует пищевую вакуоль, которая быстро втягивается в клетку и попадает в ее заднюю часть, где и происходит переваривание пищи.Сопоставление светооптических наблюдений с электронно-микроскопическими исследованиями этого процесса позволило обнаружить существенные различия в питании хоанофлагеллат и выделить 3 основных способа захвата и заглатывания пищи.
Первый характерен для одиночных видов со жгутиком. Пищевая псевдоподия образуется в основании воротничка и движется вверх по его наружной поверхности. Происходит слияние цитоплазмы псевдоподии и тентакул воротничка. При контакте с бактерией псевдоподия немного отгибается наружу, и между ее дистальной частью и воротничком образуется пищевая чашечка, в которую попадают 1—2 бактерии. Затем чашечка замыкается и образует пищеварительную вакуоль, которая быстро втягивается в цитоплазму особи.
Принципиально иным способом питаются безжгу- тиковые формы Choanoeca perplexa. Их воротничок значительно больше, чем у плавающих жгутиковых клеток, и обычно широко раскрыт. Бактерии и пищевые частицы оседают на его внутренней поверхности и постепенно концентрируются в основании воротничка. Периодически из апикальной части клетки внутри воротничка появляются пищевые псевдоподии, которые движутся вдоль его внутренней поверхности по направлению к бактериям и захватывают их в пищевую чашечку, впоследствии замыкающуюся и образующую пищевую вакуоль.
Третий способ питания характерен в основном для колониальных хоанофлагеллат. Пищевая псевдоподия образуется на уровне середины тела и по мере движения вперед и в сторону растягивает тонкую оболочку на переднем конце. В результате в образовавшееся между оболочкой и поверхностью шейки пространство втягиваются бактерии с нижней части воротничка. Пищевая псевдоподия охватывает пищу и, замыкаясь, образует у основания воротничка пищевую вакуоль. Последняя втягивается в шейку клетки и транспортируется в заднюю половину особи. При таком способе питания может захватываться по нескольку бактерий одновременно, поэтому образуются крупные пищевые вакуоли.
Размножение и жизненные формы
Бесполое размножение хоанофлагеллат осуществляется путем продольного деления клетки пополам. Описанные в прошлом веке образование спор, конъюгация и поперечное деление клеток в дальнейшем не были обнаружены.Процесс деления изучен наиболее детально у Мо- nosiga ovata (Карпов, Мыльников, 1993; Карпов, Ледбите, 1997). Базальный аппарат жгутика удваивается, а сам жгутик быстро втягивается внутрь клетки, где происходит разборка аксонемы. Шейка особи набухает, увеличиваясь в диаметре. Раздвигаются в стороны основания тентакул воротничка, и последний, если смотреть на клетку сверху, превращается из окружности в эллипс. Обе пары кинетосом мигрируют в противоположные стороны к будущим полюсам делящегося ядра, сохраняя при этом корешки из микротрубочек. Ядро вытягивается в направлении, перпендикулярном продольной оси клетки. Внутри него образуется микротрубочковое веретено, которое направляет хромосомный материал к середине ядра. На стадии поздней профазы кинетосомы находятся на полюсах ядра и уже сформированы новые короткие жгутики, ядерная оболочка еще сохраняется, а митохондрия вплотную прилегает к поверхности ядра. На стадии метафазы ядерная оболочка прорывается на полюсах ядра и микротрубочки веретена соединяются с кинетосомами. После этого начинается процесс расхождения хромосом и дальнейшее разрушение ядерной оболочки. В анафазе и ранней телофазе ядерная оболочка отсутствует и восстанавливается только в поздней телофазе. Хромосомный материал мигрирует вместе с остатками микротрубочек вслед за кинетосомами, которые постепенно перемещаются к середине апикальной части клетки. Ветви митохондрии окружают ядерный материал на всем протяжении митоза, а в анафазе и телофазе частично выполняют функции ядерной оболочки. Цитокинез завершается почти сразу после формирования дочерних ядер, а жгутик вырастает до нормальных размеров только после окончательного отделения клеток друг от друга и перемещения кинетиды в центр воротничка.
Таким образом, метафазная фигура ядра у М. ovata относится к типу ортомитоза, который за период метафазы трансформируется из закрытого в полузакрытый (по классификации И. Б. Райкова, — см.: Райков, 1982). Похожим образом протекает митоз у морского воротничкового жгутиконосца Stephanoeca diplocostata (Leadbeater, 1994).
Деление клетки S. diplocostata связано с формированием домика у дочерней особи (Leadbeater, 1994). Образование домика происходит в 2 этапа: сначала внутри клетки синтезируются и накапливаются кремниевые полоски, а затем они группами выделяются наружу и осуществляется сборка домика. Полоски синтезируются в так называемых пузырьках накопления, которые ассоциированы с микротрубочками корешковой системы. По мере «роста» полосок пузырьки вытягиваются вдоль микротрубочек, служащих опорным и направляющим образованием. На заключительной стадии формирования полоски собираются группами по 5—8 штук на периферии клетки.
На втором этапе полоски выделяются наружу и скапливаются в районе воротничка. Причем пакеты полосок будущих продольных и поперечных ребер расположены взаимно перпендикулярно. В дальнейшем, по мере процесса деления, полоски претерпевают реорганизацию при помощи тентакул воротничка. Концы полосок каким-то образом приклеиваются друг к другу, и корзинка приобретает окончательную форму взрослой особи.
Среди представителей лорикатных морских хоанофлагеллат (сем. Acanthoecidae) принято различать 2 типа деления клетки: нудиформное (nudiform) и тектиформное (tectiform). В результате нудиформного деления образуется дочерняя клетка без домика, которая уплывает в поисках подходящего субстрата для прикрепления. Такие клетки формируют домик только после прикрепления к субстрату. У тектифор- мных видов дочерняя клетка строит свой домик сразу после деления и только после этого уплывает на поиски нового места для прикрепления.
Одним из эффективных способов увеличения численности колониальных видов является деление целых колоний. Лентовидные колонии распадаются на группы из нескольких особей. При этом нет какого-либо критического количества клеток, с которого начинается фрагментация. Одной из возможных причин деления колонии может быть механическое воздействие. Однако таким образом трудно объяснить деление шаровидных колоний. Сначала колония принимает форму эллипсоида. Затем по его малому экватору возникает перетяжка, которая постепенно увеличивается и приводит к формированию 2 дочерних колоний. Деление происходит независимо от размеров, и при этом часто образуются 2 неравные колонии.
Интересно сопоставить между собой размножение и расселение хоанофлагеллат. У одиночных жгутиконосцев эти процессы протекают одновременно, т. е. деление прикрепленной особи на 2 вызывает образование расселительной формы. Для колониальных хоанофлагеллат характерно то, что расселительная форма появляется не в результате деления, как у одиночных, а более или менее независимо от этого процесса. Из колоний выходят (отрываются), как правило, взрослые зооиды, морфологически неотличимые от других особей. Расселительные формы очень важны для прикрепленных колоний — таких как Соdonosiga. У них имеется несколько одноклеточных стадий с расселительной функцией. Для плавающих колоний расселение не столь актуально, и разнообразие одноклеточных форм у них невелико.
Все изученные воротничковые жгутиконосцы образуют в своем жизненном цикле свободноплавающие особи, лишенные домиков и служащие для расселения. Для них характерны тентакулообразные выросты на заднем конце и активное движение в толще воды, чаще всего задним концом вперед. Только у одиночного вида Monosiga ovata клетки обычно плавают передним концом вперед, что значительно увеличивает их скорость передвижения.
Наибольшее многообразие форм обнаружено при исследовании культур прикрепленных колониальных хоанофлагеллат из рода Codonosiga. У них встречается до 3 расселительных форм и ряд стадий в развитии колоний. Эти колониальные стадии закономерно сменяют одна другую, чередуясь с одиночными, и образуют цикл развития. Вероятно, все отмеченные в культуре формы имеют место в природе и сменяются в зависимости от изменений условий окружающей среды.
Пресноводные хоанофлагеллаты могут формировать при неблагоприятных условиях округлые цисты с толстой оболочкой. По-видимому, это характерная черта всех воротничковых, хотя данных по морским видам пока нет. Оболочка цисты Desmarella moniliformis — единственного вида, у которого удалось получить данные по строению цисты, довольно толстая, состоит из фибриллярного материала, расположенного снаружи от плазмалеммы. Под плазмалеммой формируется тонкий плотный слой из микрофиламентов. Внутри клетки сохраняются обе кинетосомы, ядро, митохондрии и небольшая(по сравнению с ее'размерами в вегетативной клетке) диктиосома. Мебранные образования почти полностью отсутствуют.
Экология и распространение
Среди воротничковых жгутиконосцев чисто планктонных форм немного. К ним относятся преимущественно колониальные виды. Основная же масса хоанофлагеллат представлена эпибионтами и обрастателями, которые встречаются в планктоне и в бентосе. В большом количестве они поселяются на планктонных водорослях, высшей водной растительности, ракообразных и других гидробионтах. Например, Monosiga ovata использует в качестве субстрата даже сократимые стебельки сувоек и щупальца гидр. Пресноводные виды предпочитают водоемы мезотрофного типа с небыстрым течением. Одним из основных факторов, лимитирующих их численность, является кислород. Они, как правило, не встречаются в анаэробных условиях.Большинство хоанофлагеллат способны развиваться в широком диапазоне температур: от -4 до 35 °C с оптимумом примерно 20—25 °C. Лишь немногие виды предпочитают более низкие температуры и достигают высокой численности в зимне-весеннее время.
При наличии пищи воротничковые жгутиконосцы неприхотливы к солевому составу среды. Оптимальное для них значение pH несколько сдвинуто в щелочную сторону. Следует отметить, что в водоемах с большим содержанием минеральных взвесей при прочих благоприятных условиях они не развиваются. Это связано с особенностями питания хоанофлагеллат.
Воротничковые жгутиконосцы встречаются всесветно. Они отмечены в пресных водоемах всех континентов и климатических зон мира, в морях и океанах обоих полушарий и даже во льдах Антарктиды. Наибольший интерес представляет тот факт, что многие пресноводные виды встречаются в солоноватоводных водоемах и морях, а иногда и в супергалинных водах, где соленость достигает 83 %.
Филогения
Филогенетические взаимоотношения в отряде Choanoflagellida следует рассматривать исходя из факта, что клетки всех воротничковых имеют единую организацию. Следовательно, наиболее примитивно устроенные виды, несущие основные черты отряда, должны стоять ближе других к предковой форме. Такой наиболее простой и, очевидно, самой древней группой можно считать род Monosiga, От нее прослеживаются 2 основных направления в развитии Choanoflagellida: 1) образование свободноплавающих и прикрепленных колоний сем. Monosigidae; 2) образование форм, живущих в домиках.Эволюцию колоний от одиночного предка логично рассматривать, переходя от просто устроенных колоний к более сложным. Наиболее простыми представляются линейные колонии, характерные для рода Desmarella. Эта ветвь развития колоний от рода Monosiga кажется наиболее очевидной. Другая линия развития привела, по-видимому, к образованию табличных колоний типа Protospongia и шаровидных колоний типа Sphaeroeca. Еще одну самостоятельную ветвь венчают прикрепленные древовидные колонии Codonosiga. Весьма независимо развивались, вероятно, древовидные колонии на основе слизи, а не стебелька. К ним относится Sphaerodendron.
Второе направление эволюции хоанофлагеллат привело к появлению и усложнению домиков (теки). У видов сем. Salpingoecidae отмечено 2 основных типа теки, которые развивались независимо: трубчатые и бокаловидные. К первому относятся домики Stelexomonas и Aulomonas, а ко второму — Salpingoeca.
Совершенно особую ветвь развития представляют виды сем. Acanthoecidae. Их лорика отличается от теки Salpingoecidae по строению и химическому составу, поэтому оба семейства должны иметь общих предков на уровне «голых» форм.
Впервые связь хоанофлагеллат с многоклеточными организмами была отмечена Джеймсом-Кларком в 1868 г. (James-Clark, 1868). Он сравнил воротничковых жгутиконосцев с хоаноцитами губок, указал на большое сходство и тех и других и предположил, что эти 2 группы животных родственны между собой. Дальнейшие исследования позволили уже другим авторам сделать более смелые выводы. При описании нового рода Protospongia Кент (Kent, 1880— 1882) указывает на то, что «пропорции воротничковых клеток в большой колонии могут нарушаться» и некоторые особи принимают неопределенную амебоидную форму, переходя внутрь колонии. Сторонники гипотезы колониального происхождения Metazoa (см.: Иванов, 1968) использовали этот род как модель перехода от колониальности к дифференцировке клеток в соответствии с различными функциями, а затем и к образованию двухслойного организма. В общих чертах этот процесс мог выглядеть следующим образом. Воротничковые клетки, получив достаточное количество пищи, отбрасывали жгутик и воротничок, переходили в амебоидное состояние и переползали внутрь колонии для переваривания пищи. С течением времени дифференцировка на наружные воротничковые (эктодермальные) и внутренние амебоидные (энто- дермальные) клетки завершилась и образовался первый двухслойный организм, от которого произошли все современные многоклеточные животные.
Одним из главных аргументов в этой гипотезе было приведенное выше описание Кента. При более детальных наблюдениях за представителями рода Protospongia (=Proterospongia) оказалось, что ни амебоидных, ни других клеток, кроме воротничковых, в колониях всех 5 видов этого рода не существует. Нет амебоидных клеток и у других колониальных хоанофлагеллат. Таким образом, подобные модели не подтверждаются фактическим материалом.
Применение электронно-микроскопических методов показало, что воротничковые жгутиконосцы и хоаноциты губок обладают рядом общих черт. Общая морфология и симметрия клетки у них одинаковы. Сходно устроены тентакулы воротничка. Кристы в митохондриях имеют пластинчатую форму. Есть определенное сходство и в структуре жгутикового аппарата, хотя для более детального сравнения не хватает данных по строению жгутиков у хоаноцитов.
Близость по многим признакам не может быть случайной, поэтому многие авторы связывают происхождение губок с воротничковыми жгутиконосцами. Однако недавнее исследование хоаноцитов пресноводной губки Ephydatia fluviatilis (Карпов, Ефремова, 1994) показало, что имеются значительные отличия в строении их жгутикового аппарата. Так, в переходной зоне жгутика хоаноцитов расположена спиральная фибрилла, а не центральный филамент, как у хоанофлагеллат. Кроме того, в основании жгутика находится ямка, что характерно для инфузорий и не встречается у хоанофлагеллат. Конечно, для того чтобы сделать окончательный вывод о строении жгутика у хоаноцитов губок, необходимы исследования и других видов, особенно морских представителей. Тем не менее уже первые такие данные вызывают сомнения в близости хоанофлагеллат и губок.
Книги:
 Протисты. Руководство по зоологии. Часть 1. Алимов А.Ф. 2000
Протисты. Руководство по зоологии. Часть 1. Алимов А.Ф. 2000
 Протисты. Руководство по зоологии. Часть 2. Алимов А.Ф. 2007
Протисты. Руководство по зоологии. Часть 2. Алимов А.Ф. 2007
 Протисты. Руководство по зоологии. Часть 3. Алимов А.Ф. 2011
Протисты. Руководство по зоологии. Часть 3. Алимов А.Ф. 2011
 Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987
Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987



 Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
 Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423
Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423
 Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989
Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989