



























Описание:
Краткая характеристика. Млекопитающие — высший класс позвоночных животных, венчающий всю систему животного мира. Наиболее важными чертами общей организации этих животных служат:1) высокий уровень развития нервной системы, обеспечивающий сложные и совершенные формы приспособительного реагирования на воздействия окружающей среды и слаженную систему взаимодействия различных органов тела;
2) живородность, сочетающаяся (в отличие от других живородных позвоночных, например рыб и пресмыкающихся) с выкармливанием детенышей молоком. Она обеспечивает лучшую сохранность молодняка и возможность размножения в весьма разнообразной обстановке;
3) совершенная система терморегулирования, благодаря которой тело имеет относительно постоянную температуру, т. е. постоянство условий внутренней среды организма.
Все это способствует почти повсеместному распространению млекопитающих по Земле, при котором они заселяют все жизненные среды: воздушно-наземную, водную и почвенно-грунтовую.
В строении млекопитающих можно отметить следующее. Тело их покрыто волосами, или шерстью (есть редкие исключения вторичного характера). Кожа богата железами, имеющими разнообразное и весьма важное функциональное значение. Особенно характерны млечные (молочные) железы, на которые нет даже намеков у других позвоночных.
Нижняя челюсть состоит лишь из одной (зубной) кости. В полости среднего уха имеются три (а не одна, как у амфибий, пресмыкающихся и птиц) слуховые косточки: молоточек, наковальня и стремечко. Зубы дифференцированы на резцы, клыки и коренные; они сидят в альвеолах. Сердце, как и у птиц, четырехкамерное, с одной (левой) дугой аорты. Красные кровяные тельца без ядер, что повышает их кислородную емкость.
Ныне живущих млекопитающих насчитывают несколько более 4000 видов.
Строение млекопитающих. Внешний облик млекопитающих многообразен. Это объясняется удивительным разнообразием свойственной им жизненной обстановки — наземная поверхность,, кроны деревьев, почва, вода, воздух. Очень сильно варьируют и размеры тела — от 3,8 см при массе 1,5 г (у карликовой белозубки) до 30 м и даже более при массе около 150 т (у синего кита), что соответствует массе 30 слонов или 150 быков.
Кожа, как и у других позвоночных, состоит из двух слоев: наружного — эпидермиса и внутреннего — дермы, или собственно кожи . Эпидермис, в свою очередь, подразделяется на ряд слоев, из которых глубинный, или ростковый, состоит из живых, делящихся клеток, а наружный, или роговой, представлен отмершими (в связи с роговым перерождением) клетками, из них наиболее поверхностно расположенные постоянно слущиваются.
За счет деятельности эпидермиса возникают такие его производные: волосы, ногти, когти, копыта, рога (кроме оленьих), роговые чешуи и кожные железы.
Дерма сильно развита и состоит преимущественно из волокнистой ткани. Нижняя часть этого слоя рыхлая, и в ней откладывается и;up — это так называемая подкожная жировая клетчатка. Подкожная клетчатка особенно сильно развита у водных зверей — тюленей и китов, у которых она выполняет термоизолирующую роль и уменьшает плотность тела.
Общая толщина кожи у разных видов различна. Как правило, у наземных обитателей холодных стран, имеющих пышный волосяной покров, она меньше. Есть разница и в прочности кожи. Встречается своеобразная кожная хвостовая аутотомия у мышей, тушканчиков и особенно у сонь. Кожный хвостовой футляр у них легко обрывается и соскальзывает с хвостовых позвонков, что дает возможность схваченному за хвост зверьку уйти от врага. Такое же биологическое значение имеет и очень тонкая, непрочная и бедная кровеносными сосудами кожа зайцев.
Кожный покров млекопитающих имеет существенное терморегуляционное значение. Роль волосяного покрова понятна, но нужно еще указать и на значение кожных кровеносных сосудов. При расширении их просветов, которые регулируются нервнорефлекторными механизмами, теплоотдача резко возрастает. У некоторых видов существенное значение имеет также испарение с поверхности кожи пота, выделяемого одноименными железами.
Кожные железы у млекопитающих, в отличие от рептилий и птиц, обильны и многообразны. Потовые железы — трубчатые, глубинные их части могут иметь вид клубка. Эти железы выделяют в основном воду, в которой растворены мочевина и соли. Не у всех видов млекопитающих потовые железы развиты одинаково. Их мало у собак, кошек; у многих грызунов они есть только на лапах, в паховой области и на губах. Вовсе нет потовых желез у китообразных, ящеров и некоторых других. Сальные железы гроздевидные, протоки их открываются в волосяную сумку. Секрет этих желез смазывает поверхность эпидермиса и волосы, предохраняя их от снашивания и смачивания.
Пахучие железы представляют видоизменение сальных или потовых желез, а иногда их комбинацию.
Выделения всех кожных желез, как и других пахнущих выделений (например, полового и пищеварительного трактов, мочи, секрета специализированных желез), служат наиболее важным средством внутривидового и межвидового общения — химической сигнализацией млекопитающих. Особое значение этого типа сигнализации в большой мере определяется дальностью ее действия и длительностью сохранения сигнала. Таковой не теряет своего значения много времени после его подачи. Виды, имеющие определенные участки обитания, особи, пары, семьи метят участок запаховыми метками, которые они оставляют на приметных предметах: кочках, камнях, пнях, отдельных деревьях или просто на поверхности земли.
Родители оставляют запаховые метки на детенышах, в гнезде и на следах движения, за пределами гнезда или места нахождения детеныша, если гнездо не сооружается. Именно химическая сигнализация обеспечивает нахождение своих, а не чужих детенышей не делающими гнезд оленями, тюленями и такими норниками, как лисицы, песцы, соболи, куницы, полевки, мыши. В целом запаховая сигнализация несет решающее значение для выработки форм поведения млекопитающих.
Млечные железы возникли как видоизменение потовых желез. У низших однопроходных млекопитающих они сохраняют простое трубчатое строение, и протоки их открываются на определенном участке кожи брюшной поверхности. Сосков в этом случае нет. У сумчатых и плацентарных млечные железы гроздевидные, протоки их открываются на сосках. Расположение желез и сосков бывает различным. У летучих мышей и обезьян они располагаются на груди и сосков бывает одна пара. У большинства копытных соски расположены, как и сами железы, в паховой области.
У других зверей млечные железы и соски расположены на брюхе и груди. Число сосков в известной мере связано с числом детенышей, приносимых самкой в помете. Максимальное число сосков 24 (опоссумы из сумчатых, тенреки из насекомоядных).
Волосяной покров состоит из волос различного типа. Основными категориями их будут пуховые волосы, остевые, направляющие и чувствующие, или вибриссы. У большинства видов преимущественное развитие получают пуховые волосы, формирующие подшерсток, или пух. Однако у некоторых животных, например оленей, кабанов, многих тюленей, подшерсток редуцирован и волосяной покров состоит главным образом из остей.
Наблюдается периодическая смена волосяного покрова, или линька. У некоторых видов она бывает дважды в году — осенью и весной; таковы белки, некоторые зайцы, песцы, лисицы. У кротов бывает и третья, летняя линька. Суслики, сурки линяют один раз в году, в весенне-летнее время. У северных млекопитающих по сезонам меняется густота меха, а у некоторых и окраска. Так, у белки на 1 см2 поверхности огузка летом бывает в среднем 4200 волос, а зимой 8100. На зиму белеют горностаи, ласки, песцы, зайцы-беляки и некоторые другие.
Особую категорию волос представляют вибриссы — очень длинные жесткие волосы, выполняющие осязательную функцию. Они расположены на голове, нижней части шеи, груди, а у некоторых лазающих (например, у белки) и на брюхе. Видоизменениями волос являются щетина и иглы.
Кроме волос на коже млекопитающих бывают роговые чешуи. Наиболее сильно они развиты у ящеров. Роговые чешуйки имеются также на лапах (мышевидные грызуны) и на хвосте (мышевидные, бобр, выхухоль, некоторые сумчатые).
Роговыми образованиями будут полые рога копытных, ногти, когти, копыта. Рога оленей также представляют придатки кожи, но развиваются они из дермы, и вполне развившиеся рога состоят из костного вещества.
Мышечная система млекопитающих весьма дифференцирована в связи с разнообразием телодвижений. Есть диафрагма — куполообразная мышца, отделяющая брюшную полость от грудной и имеющая важное значение при вентиляции легких. Хорошо развита подкожная мускулатура. У ежей, ящеров и некоторых броненосцев она обеспечивает свертывание тела в клубок. Эта же мускулатура обусловливает поднятие игл у ежей и дикообразов, движение вибрисс. На лице — это мимическая мускулатура.
Череп характеризуется относительно крупными размерами мозговой коробки, что естественно связано с большим объемом головного мозга. Кости черепа срастаются поздно, и это обеспечивает увеличение головного мозга по мере роста животного.
В строении позвоночного столба характерны плоские сочленовные поверхности позвонков и четко выраженная расчлененность позвоночника на отделы: шейный, грудной, поясничный, крестцовый и хвостовой. Общее число шейных позвонков равно 7. Таким образом, длина шеи у млекопитающих, в отличие от птиц, определяется не числом позвонков, а их длиной. Исключение составляют только ленивцы и ламантины, у которых число шейных позвонков варьирует от 6 до 10.
Основой плечевого пояса служит лопатка, к которой прирастает рудиментарный коракоид, и только у однопроходных коракоид представлен самостоятельной костью. Ключица есть у видов, передние конечности которых совершают движения в разных плоскостях, например у обезьян. Виды, перемещающие эти конечности в одной плоскости, например копытные, ключиц не имеют.
Конечности млекопитающих вполне типичны для наземных позвоночных, но число пальцев варьирует от 5 до 1. Сокращение числа пальцев или рудиментация крайних из них наблюдается у быстро бегающих видов, например копытных, тушканчиков. Сравнительно медленно перемещающиеся звери, например медведи, обезьяны, опираются при хождении на всю ладонь и ступню (стопоходящие виды); быстрые бегуны, например собаки, копытные, опираются только на пальцы (пальцеходящие виды).
Укажем в этой связи максимальную скорость передвижения некоторых видов (км/ч): короткохвостая землеройка — 4, красная собака — 7, лесная мышь — 10, дикий кролик — 32—40, красная лисица — 72, гепард — 105—112, африканский слон —- 24—40, газель Гранта — 40—50.
Пищеварительный тракт характеризуется большой длиной и хорошо выраженной расчлепенностыо на отделы. Он начинается преддверием рта, расположенным между мясистыми губами и челюстями. У некоторых зверей расширение преддверия приводит к образованию больших защечных мешков (у хомяков, бурундуков, сусликов, некоторых обезьян). В ротовую полость открываются протоки слюнных желез, секрет которых не только смачивает пищу, но и химически воздействует (фермент птиалин) на крахмал, превращая его в сахар. Слюна вампиров, питающихся кровью, обладает антикоагулирующим свойством, т. е. препятствует свертыванию крови. У некоторых насекомоядных слюна ядовита и используется для умерщвления добычи.
Зубы млекопитающих дифференцированы на группы в зависимости от типа питания. Слабая дифференцировка свойственна малоспециализированным насекомоядным (бурозубкам). У зубатых китов дифференцировка зубов исчезла вторично.
Число зубов и распределение их по группам служат хорошим систематическим признаком. Для этого применяют зубную формулу, в которой группы зубов обозначают начальными буквами их латинских наименований: резцы — i (incisivi), клыки — с (canini), предкоренные — рт (ргае-molares) и истинно коренные — т (molares). Формулу пишут в виде дробей: в числителе —¦ число зубов в верхней челюсти, в знаменателе — в нижней. Для сокращения указывают число зубов в одной половине челюсти.
Желудок, обладающий многочисленными железами, имеет различный объем и внутреннее строение. Наиболее сложно устроен желудок жвачных копытных, поглощающих огромную массу малокалорийных и трудноперевариваемых кормов. У ящеров и муравьедов зубов нет, а желудок, как и у птиц, состоит из двух отделов: железистого-и мускулистого. Сходство усиливается тем, что во втором обнаруживаются умышленно проглоченные камешки, обеспечивающие перетирание пищи.
Кроме тонкого, толстого и прямого отделов кишечника некоторые млекопитающие имеют еще слепой отдел, в котором пища подвергается бактериальному сбраживанию. Особенно сильно слепая кишка развита у кормящихся грубой растительной пищей; длина ее достигает 1/3 длины кишечника. Длина толстого отдела кишечника по отношению к общей длине всего кишечника составляет (в %): у грызунов до 53, у насекомоядных до 30, у хищных до 22. Естественно, различна и общая длина кишечного тракта: у большинства летучих мышей он в 2,5 раза длиннее тела, у насекомоядных в 2,5—4,2, у хищных в 2,5—6,3, у грызунов в 5,0—12,0, у копытных в 12—30 раз.
Протоки печени и поджелудочной железы впадают в переднюю часть тонких кишок.
Легкие имеют сложное ячеистое строение. Самые мелкие легочные ходы — бронхиолы заканчиваются пузырьками —-альвеолами, в стенках которых ветвятся тончайшие кровеносные сосуды. Число альвеол даже у малоподвижных зверей (например, у ленивцев) равно 6 млн., а у весьма подвижных хищных оно достигает 300—500 млн. Механизм дыхания обусловливается изменением объема грудной клетки в результате движения межреберных мышц и диафрагмы.
Число дыхательных движений находится в зависимости от величины животного, определяющей разную интенсивность- обмена веществ. Оно составляет (в 1 мин): у лошади 8— 16, у черного медведя 15—25, у лисицы 25—40, у крысы 100—150, у мыши около 200. В указанной связи различна и интенсивность потребления кислорода (по Проссеру с соавт., 1977).
Соответственно всему сказанному теплопродукция у зверей высокая, хотя и в этом случае видна зависимость от размеров тела. Так, у землеройки она составляет 150 Дж/г в 1 ч, у кошки 12,51, у собаки 8,34, у слона 2,08 (по Шмидт — Нильсену, 1976).
Вентиляция легких не только обеспечивает газообмен, но и имеет терморегуляционное значение. При повышении температуры число дыханий увеличивается, а вместе с этим повышается и количество тепла, выводимого из организма. Так, у собаки отношение отдачи тепла при дыхании к общей его потере при температуре воздуха 8 °С составляет (в о/о) 14, при 15°С—22, при 30 С — 46.
Кровеносная система млекопитающих сходна с таковой птиц. Сердце полностью разделено на 2 предсердия и 2 желудочка; от левого желудочка отходит одна дуга аорты (но не правая, как у птиц, а левая). Дуга аорты отсылает к голове сонные артерии и, огибая сердце, тянется под позвоночным столбом, по пути направляя разветвленные сосуды к системам органов. Венозная система характеризуется отсутствием воротного кровообращения в почках, которое в печени, как и у других позвоночных, хорошо развито. В печени происходит нейтрализация токсических продуктов белкового обмена.
Размеры сердца варьируют в зависимости от величины тела, образа жизни, а в конечном счете — в связи с интенсивностью обмена веществ. Масса сердца, выраженная в процентах к общей массе тела, составляет: у африканского слона 0,4. у серой полевки 0,6, у обыкновенной землеройки 1,4.
Подобная зависимость прослеживается и в отношении частоты сокращений сердца: у быка массой 500 000 г число сокращений сердца в 1 мин 40—45, у овцы массой 50 000 г — 70—80, у собаки массой 6500 г —120, у мыши массой 25 г — 600.
Относительное количество крови и ее кислородная емкость у млекопитающих больше, чем у нижестоящих классов.
Своеобразные приспособления возникают у ныряющих зверей. Это выражается в повышении количества связывающего кислород глобина в мышцах (миоглобина) — около 50% всего глобина организма. Кроме того, у надолго погружающихся в веду видов отключается периферическое кровоснабжение и не меняется обеспечение кровью лишь мозга и сердца. При нырянии уменьшается также и частота сердцебиений, возникает так называемая брадикардия, приводящая к замедленному току крови и более полному использованию находящегося в ней кислорода. Например, у обыкновенного тюленя, находящегося в непогруженном состоянии, число сокращений сердца в 1 мин равно 150. Через 11 с после погружения оно уменьшается до 60, через 27 с — до 35 и далее устанавливается на уровне 30 в течение всего времени нахождения зверя под водой.
Кислородная емкость крови у ныряющих зверей наибольшая. Так, у утконоса она равна 24% на единицу объема, у ондатры 25, у обыкновенного дельфина 26, у кашалота 29%.
Все изложенное о явлениях газообмена и кровообращения наглядно подтверждает общепринятые положения о высоком уровне организации млекопитающих, что в значительной мере обусловлено интенсивностью обмена веществ и связанным с этим постоянством (на высоком уровне) температуры тела.
Головной мозг очень велик; особенно велики полушария переднего мозга, которые покрывают сверху промежуточный и средний мозг. Головной мозг по массе в 3—15 раз больше спинного, в то время как у рептилий масса их примерно одинакова. Большое развитие получает серая кора полушарий, в которой располагаются центры высшей нервной деятельности. Это и обусловливает сложные формы приспособительного поведения млекопитающих. Кора передних полушарий несет многочисленные борозды, наибольшее число которых наблюдается у высших млекопитающих. Мозжечок также сравнительно крупный и подразделен на несколько отделов.
Обонятельные органы характеризуются увеличенным объемом обонятельной капсулы и ее усложнением путем образования системы ответвлений — обонятельных раковин. Только у китообразных обонятельный аппарат редуцирован. Тюленям же свойственно достаточно острое обоняние.
Органы слуха у подавляющего большинства видов хорошо развиты. Они состоят из внутреннего и среднего уха, которые имеются и у предшествующих классов наземных позвоночных, а также из новоприобретений: наружного слухового прохода и ушной раковины, которая вторично исчезла у водных и обитающих в почве зверей. В полости среднего уха, которое отграничено от наружного слухового прохода барабанной перепонкой, находится не одна слуховая косточка — стремя, как у амфибий, рептилий и птиц, а еще две — молоточек (гомолог сочленовной кости нижней челюсти) и наковальня (гомолог квадратной кости). Молоточек упирается в барабанную перепонку, к нему причленена наковальня, которая, в свою очередь, сочленена со стремечком, упирающимся в окно перепончатого лабиринта (внутреннего уха). Вся эта система усиливает тонкость восприятия звуков. Кроме того, совершенство восприятия звуков обеспечивается и сильно развитой улиткой — извитым выростом перепончатого лабиринта, в котором расположено несколько тысяч тончайших волокон, резонирующих при восприятии звука.
У ряда зверей обнаружена способность к звуковой локации (эхолокации). Кроме хорошо известных в этом отношении летучих мышей такой способностью обладают китообразные (дельфины), ластоногие (тюлени), землеройки и, видимо, мышевидные грызуны. Землеройки при эхолокации издают импульсы частотой 30—60 кГц, продолжительностью 5—33 мкс. Морской лев при локации издает звуки частотой от 20 до 72 кГц, дельфины 120—200 кГц. Последние способны лоцировать косяки рыб с расстояния до 3 км.
Органы зрения не имеют принципиальных особенностей, и их значение в жизни зверей меньше, чем у птиц. На неподвижные предметы млекопитающие обращают мало внимания. Слабо развито или отсутствует у них цветное зрение. Так, кошки различают только 6 цветов, лошади — 4 цвета, крысы не различают зелено-желтый и сине-зеленый цвета. Только у высших приматов цветное зрение приближается к зрению человека.
Глаза водных зверей несколько похожи на глаза рыб: роговица уплощена, а хрусталик круглый, что свидетельствует о близорукости. У животных, ведущих подземный образ жизни, глаза рудиментарны, а у некоторых зверей (например, у слепого крота) они затянуты кожистой перепонкой.
Аккомодация по сравнению с птицами развита слабо и достигается только изменением формы хрусталика.
Характерной особенностью органов осязания является наличие осязательных волос, или вибрисс.
Почки у большинства млекопитающих бобовидной формы, с гладкой поверхностью. Только у китообразных, ластоногих и немногих других они состоят из нескольких долек. Основная в функциональном отношении часть почки — это ее наружный корковый слой, в котором располагаются извитые канальцы, начинающиеся боуменовыми капсулами, внутри которых находятся клубки кровеносных сосудов. В этих капсулах профильтровывается плазма крови, но не форменные ее элементы и белки. В почечных канальцах из фильтрата (первичной мочи) происходит обратное всасывание Сахаров и аминокислот. Почечные канальцы впадают в почечную лоханку, от которой берет начало мочеточник. Число почечных канальцев велико: у мыши 10 тыс., у кролика около 300 тыс.
Основной конечный продукт белкового обмена _у_ млекопитающих (как и у рыб, и амфибий) в отличие от рептилий и птиц не мочевая кислота, а мочевина. Так, у птиц мочевая кислота составляет 63—80%, мочевина лишь 1 — 10%; у млекопитающих соотношение обратное: мочевины содержится 68—91%, мочевой кислоты 0,1—8%. Такой тип белкового обмена у млекопитающих, несомненно, возник в связи с наличием плаценты, через которую система развивающегося яйца может получить, с одной стороны, практически в неограниченном количестве воду из кровяного русла материнского организма. С другой стороны, через плаценту, точнее систему ее кровеносных сосудов, из развивающегося эмбриона могут неограниченно выводиться токсические продукты белкового метаболизма. Напомним, что мочевина значительно токсичнее мочевой кислоты, но для выведения мочи такой тип обмена требует очень большого расхода воды.
Половые органы самца состоят из семенников, семяпровода, придаточных желез и совокупительного члена. Семенники у большинства располагаются в мошонке, сообщающейся с полостью тела через паховый канал. У однопроходных, китообразных, слонов и некоторых других семенники находятся непосредственно в полости тела. К семеннику прилегает его придаток, от которого отходит семяпровод, впадающий у корня полового члена в мочеиспускательный (семяизвергательный) канал. Перед впадением в мочеиспускательный канал семяпроводы образуют парные семенные пузыри — железы, секрет которых принимает участие в образовании жидкой части спермы и благодаря клейкой консистенции препятствует обратному вытеканию спермы из половых путей самки. Рядом с семенными пузырями располагается вторая парная придаточная железа — предстательная, протоки которой впадают в начальную часть семяизвергательного канала. Секрет этой железы представляет основу жидкости спермы, в которой плавают сперматозоиды.
Парные яичники всегда лежат в полости тела. В непосредственной близости от них располагаются воронки парных яйцеводов, куда и попадают яйца после их выхода из яичника. Верхние извитые отделы яйцеводов представляют фаллопиевы трубы, в которых происходит оплодотворение яйца. Далее идут расширенные отделы матки, впадающие в непарное влагалище (у некоторых сумчатых влагалище парное). Матка в простейшем случае парная, и во влагалище открываются два ее отверстия. У многих нижние отделы матки объединены, такая матка называется двураздельной (ряд грызунов, некоторые хищники). Слияние большей части маток приводит к образованию двурогой матки (некоторые хищники, китообразные, копытные). При полной утрате парности матка называется простой (некоторые рукокрылые, приматы).
Размер яиц у млекопитающих в большинстве случаев очень мал (0,05—0,4 мм), и только у однопроходных они в связи с наличием значительного количества желтка сравнительно велики (2,5— 4,0 мм).
При развитии зародыша в матке у подавляющего большинства млекопитающих формируется плацента. Ее нет у однопроходных, а у сумчатых опа зачаточна. Плацента возникает путем срастания двух наружных околоплодных оболочек (аллантоиса и прохориона), в результате чего формируется губчатое образование — хорион. Хорион образует выросты — ворсинки, которые внедряются или срастаются с разрыхленным эпителием матки. В этом месте происходит сплетение (но не сращение) кровеносных сосудов материнского организма и плода, что обеспечивает снабжение зародыша кислородом и питательными веществами и вынос углекислого газа и других продуктов обмена.
После родов в стенке матки на месте бывшего прикрепления плодов остаются темные, так называемые плацентарные, пятна. По их количеству можно установить число детенышей в помете.
Половой диморфизм у млекопитающих выражен слабо (преимущественно в размерах и общем складе): самцы обычно несколько крупнее самок и более крепкого телосложения. Но у усатых китов,, наоборот, самки заметно'крупнее самцов. Многие парнокопытные отличаются по рогам. Самки всех оленей, кроме северного, безроги.
У самцов баранов рога более мощные, чем у самок, которые иногда бывают безрогими. Не имеют рогов самки многих антилоп. У самцов ушастых тюленей развивается подобие гривы.
Половые особенности в окраске свойственны меньшинству видов. Видимо, это связано с меньшим развитием у млекопитающих способности к зрительным (в частности, к цветовым) восприятиям. Разнополые особи различают друг друга главным образом по запаху.
Биология млекопитающих. Млекопитающие распространены практически по всей Земле; их нет только на Антарктическом континенте, хотя у его побережья держатся тюлени и киты. В районе Северного полюса бывают белые медведи, ластоногие и китообразные (нарвалы).
Широко распространены млекопитающие и по жизненным средам. Достаточно вспомнить,, что наряду с наземными видами, которых большинство, ряд видов в той или иной мере связан с водной средой; многие активно летают по воздуху; кроме того, значительное число зверей обитает в почве, где проходит вся или большая часть их жизни. Ни один класс позвоночных не дал такого разнообразия форм, как млекопитающие.
Степень и характер взаимосвязей с жизненными средами различны. Некоторые животные обитают в весьма разнообразной обстановке. Например, обыкновенная лисица распространена в лесах, степях, пустынях и в горных районах. Волки обитают в еще более разнообразных условиях (кроме указанных зон для лисицы, волки встречаются и в тундре). Естественно, что у таких зверей отсутствуют ясно выраженные приспособления к жизни в какой-либо определенной среде.
Наряду с этим специфическая среда обитания определяет многие особенности зверей. Среди обитателей тропических лесов выделяются, например, южноамериканские ленивцы, паукообразные обезьяны, некоторые южноазиатские медведи, вся или почти вся жизнь которых проходит в кронах деревьев. Здесь они кормятся, отдыхают и размножаются. В связи с таким образом жизни у видов этой группы развились разнообразные приспособления. Медведи и куницы лазают с помощью острых когтей, лемуры и обезьяны имеют хватательные лапы. У некоторых южноамериканских обезьян, древесных муравьедов и опоссумов развит цепкий хвост. Наконец, обезьяны гиббоны совершают прыжки, предварительно маятникообразно раскачиваясь на передних конечностях. Своеобразное приспособление к пассивному планирующему полету развито у летяг и южноазиатских шерстокрылов.
Животные, обитающие в открытых пространствах, имеют различные формы и степень специализации. У копытных и некоторых хищников сравнительно острое зрение, их быстрый бег помогает при больших переходах в поисках корма и воды, при преследовании. Скорость бега у некоторых животных (км/ч): бизон — 40—45, жирафа — 45—50, дикий осел — 50—55, зебра — 60—65, газель Томсона — 75—80; а из хищных: лев — 75—80, гепард — 105—115. Жилищ или временных убежищ копытные не устраивают и рождают на поверхности земли развитых детенышей, способных уже в первые дни передвигаться вместе с матерью.
Весьма разнообразны виды прыгающих зверей, которые есть в различных систематических группах и в разных странах. Таковы пустынные азиатские тушканчики (грызуны), американские кенгуровые крысы (грызуны), африканские прыгунчики (насекомоядные), австралийские кенгуру (сумчатые). Характерная особенность этих зверей —¦ «рикошетирующий» бег: прыжки вперед и из стороны в сторону, как бы отскакивание от поверхности почвы. Длинный хвост играет роль балансира и руля. Скорость бега они развивают до 20 км/ч. От видов предшествующей группы они существенно отличаются тем, что устраивают, кроме кенгуру, постоянные убежища в виде нор. Новорожденные у всех слепые и голые.
Есть еще один тип зверей открытых пространств, условно обозначенный как «тип суслика». Это мелкие либо средней величины звери, населяющие степи и горные луга с относительно густым травостоем, поэтому у них нет необходимости, а вместе с тем и возможности быстро перемещаться в поисках корма. Они сооружают постоянные норы, в которых размножаются и отдыхают, кроме того, сооружают временные норы, в которых лишь прячутся при опасности. Тело у этих зверей вальковатое, на коротких ногах, бегают они медленно. В связи с наличием нор они рождают беспомощных, голых детенышей. Кроме сусликов к этой группе относятся сурки, хомяки, степные виды пищух.
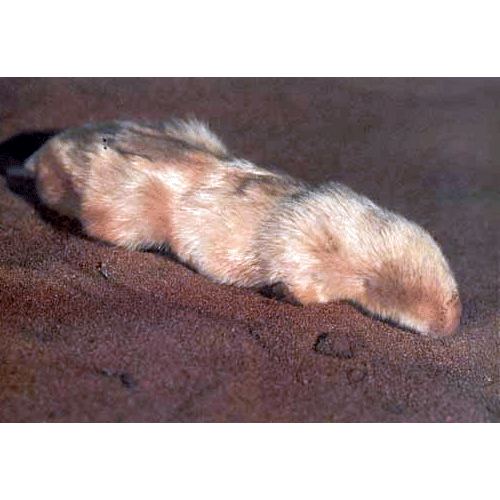
Подземные звери — высокоспециализированная, но сравнительно немногочисленная группа. Представители ее есть среди разных отрядов и во многих странах. Таковы евразийские и североамериканские кроты (насекомоядные), евразийские слепыши и слепушонки (грызуны), африканские землекопы (грызуны), австралийские сумчатые кроты. Тело у них брускообразное, без ясно выраженной шеи. Мех низкий, бархатистый. Глаза небольшие, у некоторых они скрыты под кожей. Ушных раковин нет. Хвост рудиментарный. Способ прокладывания подземных ходов у разных видов различен. Кроты роют большими, вывернутыми наружу передними лапами, вращаясь вокруг оси тела. У слепышей и слепушонок лапы слабые,, роют они резцами нижней челюсти, а лапами лишь выбрасывают землю. Система нор у этих животных очень сложная, но только небольшая часть их служит собственно жилищем, а остальная — ходами, образовавшимися при отыскивании пищи.
У водных зверей разная степень приспособления к жизни в воде. Один из видов хорьков — норка — селится всегда по берегам водоемов, где устраивает норы, но корм добывает как в воде (рыбу, земноводных), так и на суше (мелких грызунов). Существенных изменений сравнительно с наземными хорьками в ее организации нет. Более тесно связана с водой речная выдра, добывающая корм главным образом в воде (рыбу, земноводных); наземные грызуны в ее рационе играют малую роль. Плавает и ныряет она хорошо, при этом можно отметить такие приспособительные черты, как редукция ушных раковин, укорочение конечностей, наличие перепонок между пальцами.
Морская выдра, или калан, неправильно называемая морским бобром, обитает в северной части Тихого океана, корм добывает только в море (главным образом, морских ежей). На берег звери выходят для размножения, иногда для отдыха и при сильных штормах. Конечности у калана типа ластов и все пальцы объединены толстой кожистой перепонкой.
Хорошо известны полуводные грызуны: бобр, ондатра, нутрия.
Ластоногие — настоящие водные звери, но размножаются только вне водной среды — на льдах или на берегу.
Полностью водные звери — китообразные и сиреновые, способные к размножению в воде. Характеристика их дана в соответствующих местах тома. Здесь же важно подчеркнуть, что среди современных зверей можно найти весь ряд переходов от наземных через околоводных и полуводных до полностью водных форм.
Ниже приводятся сведения о глубине погружения и длительности пребывания под водой некоторых видов водных зверей (D. Davis, F. Galley, 1963).
Наконец, надо упомянуть о видах млекопитающих, перемещающихся по воздуху. И в этом случае можно выявить ряд зверей от перепрыгивающих с дерева на дерево до способных к активному полету. У белок в какой-то мере роль парашюта, определяющего начальные элементы планирования, играет распластанное при прыжке тело с вытянутыми конечностями и расчесанное надвое ©пушение хвоста. Более специализированы летяги и шерстокрылы, обладающие кожистыми складками между передними и задними лапами; у шерстокрыла складки расположены и между задними лапами и хвостом. Такое приспособление не только позволяет планировать при спуске по отлогой линии, но и служит для поворотов в воздухе и даже для незначительного поднятия. Летяги могут планировать на расстояние 30—60 м, а шерстокрылы — на 130—140 м.
К активному полету (с помощью крыльев) из млекопитающих способны только рукокрылые.
Конкретная характеристика питания разных групп приведена в соответствующих разделах. Здесь же отметим некоторые общие черты кормодобывания млекопитающими. Обращает внимание исключительное разнообразие набора пищевых объектов: почти все основные группы животных и растений. При этом особо надо подчеркнуть использование не только генеративных частей растений (семена, плоды), но и огромной массы, состоящей из вегетативных частей (стеблей, листьев, веток, коры, древесины). Значительно больше половины всех современных видов млекопитающих растительноядны, и можно утверждать, что именно появление способности использовать огромную массу растительного вещества (в тысячи раз большую, чем животного вещества) было одной из причин общего расцвета зверей.
Многие виды кормятся смешанной пищей, что отчасти связано с географическими и сезонными изменениями обилия и доступности разных объектов питания. Есть и высокоспециализированные виды, например морская выдра кормится исключительно морскими ежами. В Южной Америке есть виды летучих мышей, одни из которых питаются нектарами цветов, другие — кровью млекопитающих. Количество доедаемого корма зависит от его калорийности и большей или меньшей легкости переваривания пищи. В этой связи растительноядные виды потребляют несколько больше (по массе) пищи, чем плотоядные. Так, суточная кормовая норма (в граммах на 1 г' массы тела) выглядит следующим образом:
1) растительноядные виды: у пенсильванской серой полевки массой 46 г суточная норма составляет 0,61 г, у хлопкового хомяка массой 100 г — 0,31 г, у бобра канадского массой 12 938 г — 0,03 г, у африканского слона массой 3 672 000 г — 0,01 г;
2) плотоядные виды: у короткоухой бурозубки массой 5,4 г суточная норма составляет 1,10 г, у обыкновенной бурозубки массой 11 г — 0,62 г, у короткохвостой бурозубки массой 24,0 г — 0,53 г, у ласки массой 60 г — 0,25 г, у горностая массой 230 г — 0.21 г.
Размножение млекопитающих, характеризующееся большим разнообразием, имеет все же общие черты: внутреннее оплодотворение, живорождение (за редким исключением), выкармливание новорожденных молоком, а также устройство большинством видов специальных гнезд для деторождения.
Систематизируя основные особенности явлений размножения млекопитающих, следует выделить три основных варианта.
1. Откладывание оплодотворенного внутри тела матери «яйца» с последующим завершением его развития в гнезде (например, утконос) или в кожистой сумке родителей (например, ехидна). Яйца в этом случае относительно богаты желтком и сравнительно крупные (10—20 мм), с развитой жидкой белочной оболочкой. У ехидны одновременно созревает одно яйцо, у утконоса 1—3.
Следует оговориться, что термин «яйца» в двух приведенных выше случаях не отражает полностью существа явления. Это обусловлено тем, что у ехидны и утконоса оплодотворенные яйца задерживаются в половых путях на значительное время и проходят там большую часть своего развития. У утконоса этот период длится 15 суток, у ехидны 16 суток. Длительность же времени развития яйца вне организма матери равна соответственно 9—10 и 10—11 суткам (Гриффитси др., 1969). Таким образом, у однопроходных отмечается не настоящая яйцекладность, а скорее незавершенное яйцеживорождение.
2. Рождение недоразвитых живых детенышей,, которые развиваются в матке, но без образования настоящей плаценты. Очень слабо развитый новорожденный плотно прикрепляется к соску, который чаще открывается в полость выводковой кожистой сумки. В сумке происходит донашивание детеныша, который первоначально самостоятельно не сосет, а проглатывает молоко, впрыскиваемое ему в рот самкой. Описанный тип размножения свойствен сумчатым. Яйца у них мелкие (0,2—0,4 мм), бедные желтком, жидкая белочная оболочка развита слабо. У большинства видов одновременно развиваются единицы яиц и лишь у опоссумов — иногда более 10.
3. Рождение хорошо развитых детенышей, которые могут самостоятельно сосать молоко, а у многих видов и более или менее совершенно передвигаться. Это обусловлено полным развитием у этих видов плаценты, откуда и название описываемой группы — плацентарные млекопитающие. Яйца плацентарных очень мелкие (0,05— 0,2 мм), практически лишенные желтка, белковой оболочки нет. У большинства видов единовременно вызревает несколько яиц (до 15—18).
Длительность беременности, а в этой связи и развитость новорожденных у разных видов плацентарных зверей существенно отличны. Минимальная длительность беременности характерна для серого хомячка (11—13 суток), максимальная — для индийского слона (более 500 суток). Продолжительность внутриутробного развития зависит от совокупного действия трех обстоятельств.
Во-первых, это связано с величиной животного. Так, крошечная рыжая полевка (длина ее тела около 10 см) носит детенышей всего 18 суток, более крупная обыкновенная серая полевка (длина тела до 13 см) 20 суток, еще более крупная крыса пасюк 22 суток, хомяк (длина тела до 35 см) 30 суток, сурок-байбак (длина тела до 59 см) 40 суток, лисица (длина тела до 90 см) 52 суток.
Во-вторых, имеют значение условия деторождения, точнее, условия, в которых оказываются новорожденные. Все указанные выше млекопитающие рождают в надежно устроенных норах, и новорожденные у них голые, слепые, беспомощные.
Дикие кролики также рождают в норах, беременность у них всего 30 суток, и детеныши такие же слепые и голые. Сходной величины зайцы (мелкие расы беляка и песчаники), не устраивающие нор или гнезд, носят около 50 суток и рождают зайчат зрячих, покрытых шерстью и способных в первые же дни бегать. Близкая по размерам к сурку нутрия (длина тела до 60 см) приносит детенышей в очень примитивно устроенной норе или в наземном гнезде. Беременность у нее в 3 раза продолжительнее, чем у сурка (около 130 суток), и новорожденные развиты так же хорошо, как и у зайцев.
Велика длительность беременности у копытных, не имеющих каких-либо убежищ. У горных баранов она достигает 150 суток, у оленей и лосей 240—250 суток, у зебры 340—365 суток. Новорожденные у названных видов хорошо развиты и в первые дни после рождения уже могут следовать за матерями.
В-третьих, продолжительность беременности зависит от длительности промежутка между оплодотворением яйца и прикреплением зародыша к стенке матки, т. е. образованием плаценты. Есть виды, у которых этот промежуток очень велик,, и он в основном влияет на срок беременности. Так, у соболя и куницы начавшееся развитие яйца прерывается фазой покоя, т. е. почти полностью прекращается (латентная фаза беременности). Фаза покоя длится несколько месяцев, и в итоге у этих сравнительно мелких зверьков (длина тела около 50 см), рождающих в хорошо защищенных гнездах, беременность продолжается 230—280 суток. Детеныши рождаются слепыми и беспомощными.
Латентная фаза беременности свойственна и другим куньим (например, горностаю), а также росомахе, барсуку, тюленям, косуле и др. При этом бывает и так, что сложная беременность наблюдается только у части особей данного вида. Так, у барсуков, спаривающихся осенью, беременность длится 150 суток, а у спаривающихся летом 220—240 суток. У горностаев беременность обычно длится. 300—320 суток, и большая часть этого срока падает на латентную фазу. Но отмечаются случаи, когда латентная фаза почти или полностью выпадает и период внутриутробного развития длится лишь 42—70 суток.
В связи с условиями деторождения, различной длительностью беременности и, главным образом, с наличием или отсутствием латентной фазы сроки спаривания не имеют такого единообразия, как, например, у птиц. Волки и лисицы спариваются в конце зимы, хорьки, зайцы — весной, соболи, куницы — летом, олени — осенью. Но замечательно, что роды у подавляющего большинства приурочены к наиболее благоприятному для воспитания молодняка времени —¦ концу весны и началу лета.
Скорость размножения разных видов существенно различается. Это определяется тремя обстоятельствами: временем полового созревания, длительностью промежутка между рождениями, числом детенышей в выводке.
Вариация скорости полового созревания очень велика, что видно из следующих примеров. Рыжие полевки достигают половой зрелости в возрасте около одного месяца, ондатра не ранее 3 месяцев, зайцы, лисицы около года, соболи, волки около 2 лет, бурые медведи около 3—4 лет, моржи в 3—6 лет, зубры в 5—6 лет, слоны в 10—15 лет, носороги около 20 лет. Таким образом, этот показатель видоизменяется в пределах нескольких сотен раз.
Сильно варьирует и частота повторяемости рождений. Мелкие грызуны, например домовая мышь и обыкновенная полевка, при благоприятных температурных и кормовых условиях могут приносить в течение года до 6—8 пометов. Зайцы-русаки на юге плодятся круглый год и приносят до 4 пометов, зайцы-беляки до 3 пометов. Только один раз в году размножаются хорьки, куницы, соболи, лисицы, волки, олени и многие другие. Бурые медведи и верблюды рождают, как правило, один раз в 2 года. Один раз в 2—3 года обычно приносят приплод моржи, белые медведи, тигры, некоторые усатые киты, зубры. Этот показатель скорости размножения варьирует в десятки раз.
Существенно различна и величина выводка: плодовитость составляет от 1 до 20, чаще 12—15 детенышей. Наименее плодовиты виды, которым свойственны постоянные и подчас длительные перемещения и которые не устраивают даже примитивных временных жилищ, Таковы слоны, лошади, зебры, ослы, ламы, верблюды, жирафы, олени, тюлени, киты.
Наибольшая величина выводка свойственна мелким норным грызунам: полевкам, мышам, некоторым сусликам, У этих видов нередко бывает 10 детенышей или несколько более, в отдельных случаях даже до 20 эмбрионов. Значительна плодовитость (до 12 детенышей) некоторых зайцев, кабанов, мелких и средних норных хищников (лисица до 12, песец, горностай до 18). Крупная величина выводка у названных видов обеспечивается надежностью жилищ, в которых укрыты новорожденные, а также обилием и доступностью пищи (хотя бы в некоторые годы).
Промежуточное положение занимают крупные хищники и некоторые крупные грызуны. Пр 2—3 детеныша приносят пумы, тигры, рыси, бурые медведи. Добывать корм для молодняка этим животным труднее, чем предыдущим; большинство их не строит надежных жилищ при выводе детенышей.
Если обобщить все сказанное о скорости размножении, то можно сделать вывод, что, как правило, виды, медленно созревающие, имеют большие промежутки между рождениями и наименьшее число детенышей. Наоборот, виды млекопитающих, у которых половое созревание наступает рано, плодятся часто и выводок у них большой.
Замечено, что скорость размножения находится р тесной зависимости от продолжительности жизни; медленно размножающиеся виды живут значительно дольше высокоплодовитых. Примерная продолжительность жизни g природной обстановке такова: слона индийского 70—80 лет, зубра до 50 лет, оленя-марала до 25 лет, кита усатого около 20 лет, медведя бурого 30—50 лет, волка около 15 лет, лисицы 10—12 лет, соболя 7—8 лет, зайца 5—7 лет, полевки 10—18 месяцев. Вследствие этого скорость обновления у разных видов очень различна.
Характер брачных отношений разнообразен. Большинство видов -— полигамы: самцы оплодотворяют по нескольку самок. Таковы насекомоядные, большинство грызунов, многие хищные, копытные, китообразные. Самцы большинства полигамных видов в период спаривания не удерживают около себя самок, и в видовом строе населения не возникает в это время принципиальных особенностей (грызуны^ насекомоядные^ полигамные хищники). Самцы становятся лишь более подвижными, разыскивая неоплодотворенных самок.
Иначе протекает брачный период у ушастых тюленей и многих копытных. Котики образуют гаремы — скопления самок (иногда до нескольких десятков) около одного самца-производителя. Гарем существует более месяца, и самцы ревниво оберегают своих подруг. Самцы диких лошадей образуют косяки, самцы оленей — табуны.
Моногамия выражена по-разному. Самец настоящих тюленей обычно кроет только одну самку, но пара образуется лишь на время совокупления. Шакалы, лисицы, песцы, ондатры образуют пары на сезон размножения, и самцы у этих видов принимают участие в охране гнездового участка, а у некоторых (например, у песцов) и в воспитании молодых. Сильные самцы волков удерживают около себя самку в течение нескольких лет. Только немногие виды обезьян иногда образуют пары на всю жизнь.
Длительность молочного кормления варьирует, что связано со многими причинами, в некоторой мере с быстротой распадения выводка (см. ниже). Зайчата уже через 7—8 суток начинают есть траву, хотя одновременно сосут молоко. У ондатры лактационный период длится около месяца, у волка 4—6 недель, у песца 6—8 недель, у бурого медведя около 5 месяцев. В некоторой степени приведенные различия обусловлены питательностью материнского молока.
Время существования семьи у различных млекопитающих различно. Оно определяется в основном частотой деторождения, обилием корма и в этой связи возможностью перехода молодых к самостоятельному образу жизни, степенью трудности самостоятельного перезимовывания. Очень короткий семейный период у многоплодных мелких мышевидных грызунов, например у полевок он длится около 2—3 недель. Кормов для них обычно достаточно, и добыча их не требует особых навыков. Рано, в возрасте около одного месяца, переходят на самостоятельное существование суслики. Примерно в 2 раза дольше длится семейный период у белок, условия жизни которых более сложные. Еще дольше существует семья у хищников. Самостоятельная добыча корма в этом случае требует больших навыков. Так, у хорька семья распадается при возрасте молодых в 3 месяца, у лисицы в 3—4 месяца, у волка в 10—11 месяцев. Семьями зимуют и сурки. Почти 2 года находятся при матери бурые медведи, а тигрята 2— 3 года.
Распад семей и расселение молодняка — несомненное приспособление, предотвращающее истощение кормовых средств близ гнезда.
Приспособления млекопитающих к переживанию неблагоприятных в кормовом и погодном отношении периодов года более разнообразны и совершенны, чем у ниже стоящих классов. К зиме или засушливому лету в организме накапливаются резервные энергетические вещества, помогающие пережить тяжелый сезон. Помимо накопления гликогена в печени, многие виды в той или иной мере жиреют. Например, малый суслик весной имеет массу около 100—150 г, а в середине лета до 400 г. У сурка-байбака подкожный и внутренний жир в июне составляет 10—15 г, в июле 250—300 г, в августе 750—800 г. У некоторых особей до 25% общей массы тела составляет жир.
Сезонные приспособления выражаются и в миграциях. Осенью, по мере ухудшения кормовых условий, основная масса песцов и северных оленей откочевывает из тундры на юг, в лесотундру и даже в тайгу, где легче добыть корм из-под снега. Вслед за оленями мигрируют к югу и тундровые волки. В северных районах тундры зайцы-беляки в начале зимы предпринимают массовые кочевки на юг, весной — в обратном направлении. Горные копытные к лету поднимаются в верхние пояса гор с их богатым травостоем, зимой, по мере увеличения глубины снегового покрова, они спускаются вниз. И в этом случае наблюдаются совмещенные с копытными миграции некоторых хищников, например волков. Закономерные сезонные миграции пустынных копытных тоже зависят от сезонных изменений растительного покрова, а местами — и от характера снегового покрова. Сезонные кочевки наблюдаются у летучих мышей, китообразных и ластоногих.
В целом у млекопитающих миграции свойственны относительно меньшему числу видов, чем у птиц и рыб. В наибольшей мере они развиты у морских зверей, рукокрылых и копытных, в то время как среди видов наиболее многочисленных групп — грызунов, насекомоядных и мелких хищников — их практически нет.
Другое сезонное приспособление — это спячка, свойственная многим зверям из отрядов однопроходных, сумчатых, насекомоядных, рукокрылых, неполнозубых, хищных, грызунов. Нет зверей, впадающих в спячку, в тех отрядах, которым в наибольшей мере свойственны сезонные миграции. Таковы китообразные, ластоногие, копытные. Исключение составляют некоторые летучие мыши, у которых спячки чередуются с миграциями.
По степени глубины спячки различают Два основных варианта.
1. Сезонный сон, или факультативная спячка. В этом случае температура тела, частота дыхания в минуту и общий уровень обменных явлений снижаются мало. При изменении обстановки или беспокойстве сон легко может быть прерван. Характерен он для медведей, енотов, енотовидных собак, отчасти для барсуков. У белого медведя в берлогу залегают только беременные самки и неполовозрелые. Бурые медведи и барсуки в южных частях ареала в спячку не ложатся. О сдрторнии спящего зимой американского черного медведя представление дают такие даднре. При температуре воздуха —8 СС на поверхности кожи отмечена температура +4 °С, р прямой кишке +22 °С в ротовой полости +35 °С (против +38 °С в период бодрствования). Частота дыхания сократилась до 2—3 в минуту (против 8—14 при бодрствовании). _
Сроки залегания в зимний сон и его длительность изменчивы не только географически, но и по годам.
2. Настоящая непрерывная сезонная спячка. Она характеризуется потерей способности к терморегуляции (состояние гетеротермии), резким сокращением числа дыхательных движений и сокращений сердечной мышцы, падением общего уровня обмена веществ.
Полного прекращения обмена веществ во время спячки не бывает, и животные существуют за счет расходования энергетических запасов своего тела. В первую очередь расходуются жир и гликоген печени, а также вещества и некоторых других органов.
Наряду с зимней может быть и летняя спячка, вызванная также сезонным ухудшением кормовой базы. Чаще всего она наблюдается у грызунов, лишенных летом полноценного и богатого водой корма. Таковы в основном суслики. Наиболее рано впадает в спячку желтый, или песчаный, суслик Средней Азии (в июне — июле). У сусликов летняя спячка обычно без перерыва переходит в зимнюю.
Летняя спячка наблюдается и у обитателей тропической зоны. Сенегальские ежи спят летом 3 месяца, а примитивные мадагаскарские насекомоядные — тенреки — 4 месяца.
Новое (сравнительно с предшествующими классами) основное приспособление, обеспечивающее переживание неблагоприятных сезонных условий жизни,— собирание запасов корма. Оно в различной мере свойственно разным систематическим группам млекопитающих. Не запасают корм классические кочевники — номады: китообразные, ластоногие, летучие мыши, копытные и млекопитающие, впадающие в спячку. В зачаточной форме это наблюдается у насекомоядных. Некоторые землеройки, например североамериканские короткохвостые землеройки (Blarina), делают лишь небольшие запасы из беспозвоночных.
Более распространено захоронение излишней добычи у хищных. Ласки и горностаи собирают по 20—30 полевок и мышей, черные хори складывают подо льдом по нескольку десятков лягушек, норки — по нескольку килограммов рыбы. Более крупные хищники (куницы, росомахи, кошки, медведи) прячут остатки добычи в укромных местах, под упавшими деревьями, под камнями. Леопарды нередко укрывают часть добычи в ветвях деревьев.
Характерной особенностью запасания корма хищниками служит то, что для его захоронения не сооружается специальных кладовых, запасом пользуется только одна особь. В целом запасы служат лишь малым подспорьем для переживания малокормного периода.
По-иному запасают корм различные грызуны и пищухи, хотя и в этом случае наблюдается разная степень совершенства запасания и его значения. Летяги собирают по нескольку десятков граммов концевых веточек и сережек ольхи и березы, которые они хранят в дуплах. Белки складывают в опавшие листья, дупла и в землю желуди и орехи. Они развешивают также грибы на ветках деревьев. Одна белка в темно-хвойной тайге запасает до 150—300 грибов, а в ленточных борах Западной Сибири, где кормовые условия хуже, чем в тайге, до 1500—2000 грибов, преимущественно маслят. Сделанные белкой запасы используют многие особи этого вида.
В Восточной Сибири и на Камчатке большие запасы корневищ, клубней и семян (за год до 15 кг) делает маленький зверек полевка-экономка. Характерно, что в западных частях ареала названного вида, где зима менее продолжительная и мягкая, полевка-экономка запасов не делает. Однако много запасает на зиму корма в виде травы и луковиц лука обитатель континентальных степей Забайкалья и Северной Монголии — полевка Брандта.
Большая песчанка и пищухи запасают траву или высушенное сена, складывают их в норы, под наземные укрытия или на поверхности земли стожками. Бобры собирают на зиму запасы веточного корма, который чаще убирают в воду около гнезда, реже складывают вне воды. Находили запасы веток объемом до 20 м3.
Характерными особенностями запасания служат множественность запасов, обеспечивающих зверей кормом в голодный период, устройство специальных хранилищ для запасенного корма и коллективное, чаще семейное его употребление.
В заключение укажем, что корм запасают и немногие виды зверей, впадающих на зиму в спячку. Таковы бурундуки и сибирские длиннохвостые суслики. Собранный в местах спячки корм используется этими видами весной, когда пробудившиеся зверьки еще не обеспечены вновь появившейся пищей.
Численность видов млекопитающих по годам меняется в широких пределах. Особенно значительные изменения наблюдаются у массовых, быстро размножающихся видов. Таковы многие грызуны, зайцеобразные, некоторые хищники. Кроме того, в больших пределах изменчива численность некоторых копытных: кабана, косули, северного оленя, сайгака и некоторых других. У сравнительно редких и медленно размножающихся видов, как, например, у крупных хищников (медведи, крупные кошки), резких изменений по годам нет или они менее очевидны.
В основе непостоянства численности зверей лежат многообразные и не всегда ясные причины. Можно утверждать, что различие в обилии особей по годам возникает в результате изменения интенсивности размножения и скорости отмирания особей видов.
Одна из наиболее распространенных причин колебаний численности млекопитающих — изменчивость обилия кормов и их доступность. Периодически повторяющиеся неурожаи семян хвойных ставят белку в очень трудные условия, в результате которых сокращается размножение и повышается смертность. Подобного рода зависимость численности от изменчивости обилия кормов установлена для песца, лисицы, горностая и других зверей. Так, годовая плодовитость самки белки в кормные годы равна 15—20 молодых, а в годы неурожая семян хвойных всего 4—6. Средняя численность помета у песцов в годы массового размножения леммингов составляет 8 —12, а при резком сокращении численности последних — 3—6. Виды зверей, использующие в пищу вегетативные части растений (траву, ветки, кору), значительно реже и в меньшей мере страдают от бескормицы.
Другой важной причиной непостоянства численности служат заболевания, принимающие в определенных условиях эпизоотический характер. Показательно, что эпизоотии чаще возникают среди видов, обеспеченность которых кормами по годам существенно не меняется. Таковы многие виды зайцев, ондатра, водяная полевка и др. Природа эпизоотий разнообразна. Широко распространены гельминтозы, например легочно-глистная болезнь, заражение печени плоскими червями трематодами, болезни, вызываемые простейшими (кокцидиоз, пироплазмоз, токсоплазмоз), разнообразные бактериальные и вирусные заболевания (туляремия, псевдотуберкулез, нек-робацнлез, сибирская язва, чума плотоядных и др.).
Следует учитывать, что массовые заболевания не только приводят к непосредственной гибели зверей, но и снижают плодовитость, а также облегчают преследование жертвы хищниками и делают животных менее стойкими к пагубному воздействию неблагоприятных условий погоды.
Немаловажное значение для многих видов имеют отклонения условий погоды. Весенние заморозки с дождями и снегопадами губят ранние выводки зайцев, главным образом зайца-русака. Глубокие снега ставят в тяжелое положение многих копытных: кабанов, сайгаков, джейранов, маралов. Из хищников от глубокого снежного покрова и гололеда особенно страдают корсаки, лисицы, енотовидные собаки и др. Выпадающие иногда зимой дожди вызывают массовую гибель мышевидных грызунов, погибающих от затопления нор и невозможности достать корм из-под ледяной корки. При отсутствии снегового покрова или недостаточной его мощности сильные морозы приводят к гибели в промерзшей почве кротов и землероек.
Значение хищников в изменениях численности жертв различно для разных видов. Зайцы-беляки, бурундуки, водяные полевки хотя и страдают ощутимо от хищников, все же хищники обычно лишь усиливают скорость отмирания, которая в основе вызвана другими причинами. Однако в отдельных случаях могут возникать такие ситуации, в которых высокая численность хищников при низкой численности жертв может надолго задержать восстановление последних. Для медленно размножающихся копытных урон, наносимый хищниками, как правило, бывает большим, чем для многоплодных грызунов.
В недавнее время установлены внутрипопуляцпонные механизмы регуляции численности. Суть их в снижении размножаемости при резком возрастании численности (плотности популяции). Внешне это выражается в замедлении скорости полового созревания, увеличении доли прохолоставших самок (особенно молодых), сокращении длительности сезона размножения. Ф. Б. Ч е р н я в-с к и й (1982) установил, что резкому нарастанию численности копытного лемминга на о-ве Врангеля предшествует его интенсивное размножение еще под снегом. Летом доля народившихся в такой обстановке достигает 50% всего поголовья; при спаде численности и низком ее уровне этот показатель составляет около 15%. Дикий кролик в Северном Уэльсе при низкой его численности размножается почти круглогодично, а средняя величина помета достигает 6,8; наоборот, в годы высокой численности размножается только в марте — начале июня, а средняя величина выводка равнялась лишь 4,1. В ньюфаундлендском стаде гренландского тюленя при высокой численности зверей к шестилетнему возрасту созревало 50% самок и только к восьмилетнему все 100%. При сильно разреженной промыслом численности уже к 4 годам созревало 50% самок, а к 6 годам все 100%.
Причины описанных явлений выяснены недостаточно. Полагают, что большая плотность населения усиливает беспокойство особей, вызванное повышенным восприятием звуковых, запаховых, зрительных сигналов о присутствии друг друга. Усиливается конкуренция за убежища и участки обитания, отчасти и за корм. У ряда видов усиливается агрессивность поведения. Все это, воспринятое органами чувств, передается через нервную систему не только организму в целом, но и таким важным в регуляции размножения железам внутренней секреции, как гипофиз, надпочечники, гонады, зобная железа. Изменение эндокринного баланса, видимо, и служит одной из важнейших причин изменений интенсивности размножения и быстроты отмирания.
Динамика численности млекопитающих проявляется во времени и пространстве с известной закономерностью. У ряда видов она имеет некоторую, хотя и не строго выдержанную, периодичность. У песца цикл изменения численности обычно равен 3—4 годам, у таежного зайца-беляка 10—12 годам, В южных частях ареала последнего вида подъемы и спады бывают чаще, а интервалы между ними менее определенны. Кроме того, здесь меньше размах колебаний. Изменение численности вида в ту или иную сторону охватывает одновременно не всю площадь ареала, а лишь отдельные ее части. Пределы пространственного распространения этого явления в большей мере зависят от степени разнообразия, мозаичности природных условий: чем они однотипнее, тем на больших пространствах наблюдаются сходные изменения. Наоборот, в условиях разнообразного ландшафта распределение численности имеет весьма пестрый характер.
Практическое значение млекопитающих. Значение млекопитающих в жизни человеческого общества весьма разнообразно. Это определяется, с одной стороны, обилием и разнообразием видов зверей, с другой — разнообразием хозяйственной деятельности человека. В целом класс млекопитающих представляет собой огромное природное богатство, заслуживающее бережного отношения. При этом надо учитывать разное значение некоторых видов в различной природной и хозяйственной обстановке.
Так, вблизи посевов и при закладке защитных лесных полос несомненно вредны суслики, полевки и мыши. Вместе с тем ими питаются пушные хищники, мех которых — большая товарная ценность. В таежной полосе Восточной Сибири суслик (длиннохвостый) и полевки не причиняют вреда сельскому хозяйству; полевки составляют основу кормового благополучия для таких, ценных зверей, как соболь и колонок. Лисица и хорьки нежелательны близ птицеферм и на территории охотничьих хозяйств, где разводят фазанов и других птиц как объект охоты.
Некоторые грызуны имеют существенное эпидемическое значение, так как являются хранителями и передатчиками опасных для человека болезней. Кроме того, они служат прокормителями клещей и некоторых насекомых, распространяющих эти заболевания, называемые антпрогозоонозными. Сурки, суслики, песчанки, крысы — бациллоносители и распространители страшного заболевания людей — чумы. Чумные палочки передаются человеку при непосредственном контакте или через питающихся на больных зверьках блох. Микробоносителями туляремии среди млекопитающих служат преимущественно грызуны, в первую очередь водяная полевка, обыкновенная полевка,; домовая мышь, суслики, зайцы. Возбудитель передается человеку через кровососущих насекомых, клещей, через воду или при непосредственном контакте с больным зверьком, например при снятии с него шкурки. При эпизоотиях пастбищных сыпнотифозных лихорадок возбудители (риккет-сии) передаются от больных грызунов клещами. При вирусных энцефалитах (тяжелое поражение центральной нервной системы) возбудителей от больных зверьков также передают клещи.
С грызунами — вредителями сельского хозяйства или опасными в эпидемическом отношении — ведется планомерная борьба.
Диких млекопитающих добывают Для получения разнообразных продуктов, а также ведут на них спортивную охоту, которая до настоящего времени не потеряла значения.
По промыслу диких зверей СССР занимает первое место в мире. Примерно из 350 видов, свойственных нашей фауне, около 150 видов потенциально могут служить объектами добывания при промысловой и спортивной охоте или для отлова в целях расселения и содержания в зоопарках и лесопарках. Больше всего таких видов в отрядах грызунов (около 35), хищных (41), парнокопытных (20), ластоногих (13), насекомоядных (5), зайцеобразных (5—8).
Для получения пушнины добывается около 50 видов диких зверей, но основу пушной добычи составляют примерно 20 видов. Среднее долевое значение (по стоимости) наиболее важных видов видно из следующих данных.
В конкретные годы долевое значение зверей несколько меняется в связи с изменениями их численности в природе.
Кроме пушного промысла в нашей стране широко развито добывание копытных. Ежегодно их отстреливается около 500—600 тыс. голов. Товарный выход мяса при этом исчисляется цифрой около 20 тыс. т. Кроме того, получают много кожи и лекарственного сырья (пантов оленей, рогов сайгака). Добыча копытных ведется организованно, по специальным разрешениям. Годовая добыча лосей в целом по СССР оценивается в 60 тыс. голов, кабана и косули примерно по 50 тыс. голов; размеры добычи сайгака весьма изменчивы но годам.
Для ведения комплексного охотничьего хозяйства в нашей стране организованы многочисленные спортивные и промысловые хозяйства, за которыми закрепляются охотничьи угодья. Площадь таких угодий только в РСФСР составляет около 100 млн. га.
В промысле ластоногих наибольшее значение имеет добыча котиков и некоторых видов настоящих тюленей. Первых в настоящее время добывают преимущественно на о-вах Берингова моря (Командорские — СССР, Прибылова — США) и на о-ве Тюленьем в Охотском море (СССР). Добыча осуществляется на береговых лежбищах в начале лета. В Южном полушарии чрезмерный промысел южных котиков был развит в XVIII— XIX вв. Теперь во всех районах зверь полностью исчез. В современных условиях добыча южных котиков ведется местами на юге Африки, у берегов Южной Америки и ряда островов Мирового океана. Некоторые виды южного котика занесены в Красную книгу MCOII.
Из числа настоящих тюленей наибольшее промысловое значение имеют гренландский тюлень и хохлач, которых добывают на ледовых залежках в Северной и Северо-Западной Атлантике у о-вов Ян-Майен, Исландии, Ньюфаундленд и в северной части Белого моря, где ведут весьма ограниченный промысел только гренландского тюленя.
Местами в субантарктических районах Мирового океана, на береговых лежбищах, добывают морских слонов. В северной части Тихого океана регулярно промышляют несколько видов тюленя: кольчатую нерпу, полосатого и обыкновенного тюленей. Используют жир, шкуры. У некоторых видов, например у гренландского, каспийского, новорожденные имеют густой белый мех и шкурки их используют как пушнину.
Китобойный промысел в настоящее время резко сокращен на основе международных соглашений. В Южном полушарии почти полностью запрещена добыча китов в открытых океанических водах. В Северном полушарии разрешен очень ограниченный промысел немногих видов.
Актуальной задачей современности следует считать охрану животного мира, в частности млекопитающих. По данным Международного союза охраны природы, только за четыре последних столетия исчезло 62 вида зверей.
При учете видов зверей, исчезнувших с 1600 г., Фишер с соавторами (1976) выявил, что основной причиной исчезновения были: охота (33%), естественные причины (25%), вселение хищников (17%), вселение прочих животных (6%), разрушение биотопов (19%).
В Красную книгу МСОП, опубликованную в 1979 г., внесено 226 видов и 79 подвидов исчезающих и редких зверей. В их числе копытные (29 видов), низшие обезьяны (17), все виды человекообразных обезьян, выдры (8) и др. В Красную книгу СССР (1984) включен 91 вид и подвид редких и находящихся под угрозой исчезновения млекопитающих. Приведем несколько примеров.
Выхухоль — эндемик нашей фауны, спорадически распространенный в бассейнах Волги, Дона и Урала.
Амурский и туранский подвиды тигра. Первый сохранился в числе около 150 штук в Приморском крае. Второй, ранее обычный по течениям Аму-дарьи, Сырдарьи, Или и других рек, в настоящее время в СССР регулярно не встречается. Иногда заходит из Ирана и Афганистана.
Снежный барс — очень редкий вид высокогорий Средней Азии и Казахстана, частью Западной Сибири.
Гепард ранее был широко распространен в пустынях Средней Азии; в последние годы в СССР не обнаружен.
Тюлень-монах, в прошлом изредка встречавшийся у берегов Крыма, в наши воды очень редко заходит из прибрежных вод Турции и Балканского п-ова.
Из числа усатых китов в Красную книгу СССР внесено 5 видов, среди них особенно редки гренландский и северный синий киты.
Кулаи, ранее широко распространенный в Средней Азии и Казахстане, сохранился у нас только в Бадхызском заповеднике (юг Туркмении). Был акклиматизирован на о-ве Варсакельмес (Аральское море), откуда переселен в Прибалхашье.
Горал сохранился только в южной части хребта Сихотэ-Алинь (Приморский край). Общая численность около 600 голов.
Винторогий козел — также очень редкий вид, сохранившийся у нас в горах верховьев Аму-дарьи и Пянджа.
Закаспийский, туркменский и бухарский горные бараны в крайне ограниченном числе сохранились местами в горах юга Туркмении и в Таджикистане.
Воздействие на фауну осуществляется не только благодаря охране отдельных видов или целых природных комплексов, но и вследствие обогащения фауны новыми видами. Разрозненные попытки искусственного расселения млекопитающих предпринимались в России еще несколько веков назад. Существенного эффекта они не давали. Научное обоснование работ по акклиматизации дал в 1856 г. профессор А. П. Богданов. Но только после Великой Октябрьской социалистической революции работы по акклиматизации и реакклиматизации начали проводить планово и в больших масштабах. Идейным и практическим руководителем работ был профессор Московского университета Б.М. Житков. С 1925 по 1972 г. в охотничьих угодьях СССР выпущено для расселения около 500 тыс. зверей, принадлежащих к 45 видам.
Наибольшее число видов (6) завезено из Северной Америки: ондатра, енот-полоскун, американская норка, скунс, овцебык. Южноамериканское происхождение имеют 2 вида (нутрия и шиншилла), западноевропейское происхождение — 2 вида (лань и кролик). Все остальные обитают в Советском Союзе.
Акклиматизация чужеземных видов зверей проводилась и в других странах. В Западной Европе широко расселена лань, местами распространены корсиканский муфлон и ондатра. В Англии была акклиматизирована серая американская белка, которая вскоре настолько размножилась, что стала вредить хозяйству, в результате чего ее начали истреблять. В Северную Америку завезены заяц-русак, кабан, благородный олень, пятнистый олень, индийские олени замбар и аксис, индийская антилопа нильгау. Были случаи побега с ферм разводимых там муфлонов, африканских антилоп гну и зебр. Местами эти виды теперь существуют в природной обстановке.
Около 20 видов зверей (чаще преднамеренно, реже случайно) завезено в Новую Зеландию, где местных млекопитающих практически нет. Умышленно акклиматизированы 3 вида австралийских сумчатых (чернохвостый кенгуру и 2 вида кузу), благородный олень, американский олень вапити, лань, индийский олень замбар, американский лось, гималайский козел тар, серна, хорек, горностай,, ласка, кролик, заяц-русак, еж. Случайно завезены домовая мышь и 2 вида крыс. Общий облик новозеландской фауны после акклиматизации зверей и птиц (24 вида) и одичания некоторых домашних животных, например свиней, изменился до неузнаваемости.
Известны случаи, когда завоз п акклиматизация зверей приносили вред. Так обстоит дело с акклиматизацией европейского кролика в Австралии, где он оказался вредителем сельского, главным образом пастбищного, хозяйства. Кроликов не могли истребить завезенные сюда и акклиматизированные хищники (лисицы, хорьки) и даже распространение среди кроликов особо опасной для них болезни — миксоматоза. Поэтому к проблеме акклиматизации зверей надо относиться с большой, осторожностью и вести ее под строгим контролем.
Приручение и одомашнивание млекопитающих, начатое в глубокой древности, продолжается и сейчас, хотя виды зверей находятся на разной степени одомашнивания и приручения. Полностью одомашненными надо считать те виды, которые хорошо подчиняются человеку и свободно размножаются в неволе. Таковы, например, собака, лошадь, рогатый скот и многие другие. Однако бывает и так, когда указанные два условия не совпадают. Тысячелетия назад были приручены слоны, послушно выполняющие разнообразные работы. Но в неволе слоны размножаются очень редко. Львы и тигры хорошо размножаются в неволе, но не приручаются к человеку и не могут считаться одомашненными животными.
В малой мере одомашнены некоторые пушные звери, хотя в клетках они успешно размножаются. Таковы песцы, лисицы, норки, соболи и некоторые другие.
Пушное звероводство возникло менее ста лет назад, и число видов, разводимых в неволе, продолжает увеличиваться. Сравнительно недавно стали разводить норок и шиншилл. Однако, зная законы наследования признаков, в результате направленного подбора и отбора за короткий срок удалось вывести ряд новых пород. Таковы, например, серебристо-черная лисица и разнообразные цветные породы норок.
По сравнению с дикими предками мало изменились одомашненный северный олень, верблюд, як и южноазиатские быки. Это обусловлено тем, что условия содержания их мало отличаются от обстановки, в которой живут дикие родичи. Кроме того, не регулируется в должной мере подбор пар и отбор потомства. Наоборот, большинство домашних'зверей образует значительное разнообразие пород, отличных не только по внешнему виду, но и по хозяйственному использованию. Это определилось в основном селекционной работой, при которой человек отбирал животных по каким-либо нужным для хозяйства признакам. Так возникли многочисленные породы лошадей, коров, овец, собак.
Нужно учитывать, что некоторые домашние животные возникли, видимо, не от одного, а от нескольких диких предков. Ряд ученых полагают, что одни породы собак произошли от волка, другие — от шакалов. Так же обстоит, вероятно, и с происхождением пород овец.
В целом история одомашнивания и породообразования во многом еще остается невыясненной и предстоят многообразные палеонтологические, археологические и генетические исследования. Вместе с тем несомненны большие перспективы приручения и одомашнивания новых видов млекопитающих с разнообразными хозяйственными и эстетическими целями. Наряду с акклиматизацией диких зверей это обеспечит увеличение разнообразия видов.
Происхождение млекопитающих. Предками млекопитающих, несомненно, были древние палеозойские рептилии, еще не утратившие некоторых черт строения амфибий: кожные железы, слабое ороговение эпидермиса кожи, отсутствие или слабое развитие роговых чешуй, малые размеры полушарий переднего отдела головного мозга и некоторые другие, более специальные особенности. Такой предковой группой считают териодонтов, или зверозубых (Theriodontia),— подотряд терапсидных рептилий, существовавший со второй половины пермского до середины юрского периода, т. е. с конца палеозойской до середины мезозойской эры.
Прогрессивным териодонтам, жившим в начале мезозоя (триас), было свойственно вторичное костное нёбо, подобное таковому млекопитающих. Оно укрепляло верхнечелюстные кости и служило своего рода помостом, на котором заклыковые зубы дробили твердые части пищи, например хитиновые покровы членистоногих. Кроме того, костное нёбо делало возможным задерживание комка пищи во рту, не прерывая при этом дыхания. Заклыковые зубы (гомологичные коренным зубам) помимо нескольких основных вершин имели на коронках придаточные бугорки, увеличивающие эффект измельчения пищи. Имелись щечные железы, которые предположительно можно рассматривать как гомологичные слюнным Железам млекопитающих.
Конечности располагались в значительной мере под телом: плечо под углом в 45°, бедро — 55°. В плечевом поясе характерна частичная редукция коракоида (у настоящих зверей он сохраняется лишь в виде отростка лопатки). Следует указать также на разрастание подвздошных костей таза и увеличение числа крестцовых позвонков до пяти. Все эти особенности, несомненно, увеличивали возможности двигательной активности. Обособление поясничного отдела позвоночника позволяет предполагать наличие диафрагмы и соответственно «маммального» механизма вентиляции легких.
У некоторых териодонтов на костях челюстей обнаружены следы прикрепления губных мышц. Образование мягких губ служило важной предпосылкой формирования сосущего ротового аппарата, столь необходимого при молочном кормлении. Следует учесть также значительное увеличение обонятельных капсул, длина которых достигала почти половины длины черепа. Напомним в этой связи исключительно важное значение обоняния в жизни млекопитающих. Наконец, есть основание предполагать наличие у многих териодонтов роговых волосовидных образований, расположенных по наружному краю челюстей, которым, видимо, была свойственна осязательная функция. Они и обозначаются как «протовибриссы» (JI. П. Татарине в, 1976). Следовательно, и чувствующая система у териодонтов действовала совершенно.
Морфологически наиболее прогрессивными, близкими к млекопитающим зверозубыми были цинодонты (Cynodontia). Среди разнообразных триасовых цинодонтов известно много форм, близких по отдельным признакам к примитивным млекопитающим.
Древнейшие млекопитающие известны из позднего триаса Европы, Китая и Южной Африки. Несмотря на фрагментарность их ископаемых находок, очевидно, что в то время, т. е. около 200 млн. лет назад, уже существовали группы, принадлежавшие к трем различным эволюционным ветвям млекопитающих. Не исключено, что они имеют самостоятельное происхождение от различных цинодонтов. С одной из этих ветвей связаны так называемые многобугорчатые млекопитающие (отряд Multituberculata), достигшие расцвета в конце мела — начале палеогена и вымершие в позднем палеогене. Строение их черепа и зубов, особенно резцов и коренных, позволяет думать, что эти мелкие зверьки были первыми растительноядными млекопитающими, экологически сходными с грызунами. Впрочем, из мезозойских млекопитающих они были самыми крупными (некоторые достигали размеров сурка). Представители второй ветви — триконодонтов (отряд Triconodonta), вымерших в конце мелового периода, по размерам были не крупнее крысы, а по способу питания — плотоядными и насекомоядными.
Многобугорчатых и триконодонтов часто сближают с самыми примитивными из ныне живущих зверей — однопроходными (Monotremata), объединяя эти три группы в подкласс первозверей (Prototheria) и противопоставляя их настоящим (териевым) млекопитающим (подкласс Theria). Скудость палеонтологических данных по однопроходным не позволяет однозначно решить вопрос об их родстве с той или иной группой мезозойских млекопитающих. Отнесение к Monotremata единичных зубов териевого строения, найденных в миоценовых, а недавно и в р а пне меловых отложениях Австралии, позволило некоторым палеонтологам ставить вопрос о большей близости однопроходных к настоящим зверям. Вопрос этот остается дискуссионным, однако своеобразие организации Monotremata по сравнению с остальными современными млекопитающими не вызывает сомнения в их значительной обособленности от Theria и независимой эволюции по крайней мере с середины мезозоя.
Происхождение настоящих зверей (сумчатых и плацентарных) связывается с эволюцией третьей ветви триасовых млекопитающих, так называемых симметродонтов (отряд Symmetrodonta). Вместе с их вероятными юрскими потомками эупантоте-рнями эти млекопитающие составляют группу пантотериее (Pantotheria), рассматриваемую обычно в качестве инфракласса, входящего в состав подкласса настоящих зверей. Хотя пантотерин (особенно ранние симметродонты) имели многие архаичные черты строения черепа, восходящие к рептилиям, у них постепенно формируется трех-бугорчатый (или бугорчато-режущий) тип задне-коренных зубов, который принимается за исходный для всех более поздних настоящих млекопитающих (ранее всех или некоторых пантотериев называли «трехбугорчатыми»).
В поздней юре среди эупантотериев появляются формы, у которых число заклыковых зубов редуцируется от 11 до 8 и приближается к таковому у сумчатых п плацентарных. Пантотерии были мелкими зверьками, размером не более крысы. По образу питания они были близки к насекомоядным, но судя по строению зубов могли расширять свой пищевой рацион за счет как животной, так и растительной пищи. Недавно в верхнеюрских отложениях Португалии впервые обнаружен целый скелет млекопитающего, отнесенного к пантотериям. По размерам и пропорциям тела этот зверек, существовавший около 140 млн. лет назад и имевший длину тела около 20 см, напоминал тупайя. У него были укороченные передние конечности, длинный хвост, а в области таза присутствовали сумчатые кости.
В начале мелового периода бугорчато-режущее строение коренных зубов некоторых потомков пантотериев еще более приблизилось к настоящим зверям, как, например, у эзиалодона (Aegialodon) из нижнего мела Англии. Предполагается, что сумчатые (Metatheria) и плацентарные (Eutheria) произошли от форм такого типа, родственных пантотериям, хотя точное время расхождения этих Двух основных стволов современных млекопитающих недостаточно ясно. Если существование настоящих плацентарных в раннемеловое время (Ааия) и на рубеже раннего и позднего мела (Северная Америка) остается предметом дискуссий, то на протяжении позднего мела несомненные Eutheria появляются уже не только на этих материках, но и в Южной Америке и, вероятно, в Европе. К концу мезозойской эры (около 65 Млн. лет назад), т. е. ко времени окончательного исчезновения динозавров, и сумчатые и особенно плацентарные млекопитающие уже достигли значительного разнообразия и были широко расселены по земному шару.
Сумчатые млекопитающие к этому времени жили в Северной и Южной Америке и, очевидно, в Австралии, Хотя известные их находки на этом материке не древнее олигоцена. В палеогене отдельные находки сумчатых известны на всех материках, включая Антарктиду, Африку и Азию (недавно зуб сумчатого найден в олигоцене Казахстана), однако основная радиация этой группы происходила в Австралии и Южной Америке. Оба эти материка на Протяжении большей части кайнозойской эры были изолированы от других масс суши. В Австралии на протяжении многих миллионов лет вообще не было плацентарных млекопитающих, и сумчатые достигли здесь максимального разнообразия (до 18 вымерших и современных семейств), освоив многие экологические ниши, занятые на других материках плацентарными. В Южной Америке сумчатые, по всей вероятности, с самого начала жили вместе с плацентарными, но последние были представлены весьма ограниченным набором групп (без хищников), поэтому радиация сумчатых и на этом материке была исключительно широкой (от 8 до 12 кайнозойских семейств, из которых до наших дней сохранились всего 2). Мезозойские (меловые) плацентарные млекопитающие изучены пока недостаточно. Уже в меловом периоде они разделились на несколько самостоятельных групп, послуживших основой широчайшего расцвета плацентарных в палеогене и давших начало главным ветвям, от которых происходят все современные отряды. Однако непосредственные связи этих ветвей друг с другом и последовательность их отделения от общего ствола плацентарных далеко не ясны. Многие представители разных групп мезозойских плацентарных по образу питания были близки к насекомоядным. Долгое время их относили к отряду насекомоядных (Insectivora), считая его исходным для всех остальных отрядов плацентарных. Сейчас эти примитивные меловые-ранне-палеоценовые группы с неясными родственными связями иногда объединяют под названием прото-плацентарных (Proteutheria). Вместе с тем в конце мелового периода уже, по-видимому, существовали предки настоящих насекомоядных, происходившие, по всей вероятности, от различных протоплацентарных. Появляются к концу мелового периода и первые представители отрядов млекопитающих, обнаруживающих тенденцию перехода к растительной пище (пантодонты, кондиляртры). Оба отряда достигли расцвета в палеогене и впоследствии вымерли, но с кондиляртрами связано происхождение нескольких современных отрядов млекопитающих.
Морфологические данные указывают на большую примитивность неполнозубых (отряд Edentata) по сравнению с остальными плацентарными. Это подтверждают и современные данные молекулярной биологии. Поэтому можно думать, что и ветвь неполнозубых обособилась еще в меловом периоде, хотя их ископаемые остатки известны лишь с позднего палеоцена. Развитие этого отряда происходило в- основном в Южной Америке, но недавно обнаружено, что в палеогене представители неполнозубых жили и в Европе, и в Азии.
Палеоцен, начальная эпоха палеогенового периода — время бурного расцвета плацентарных млекопитающих. В это время (55—65 млн. лет назад) уже известны многие отряды, сохранившиеся до наших дней. От одной из групп «протоплацентарных» — позднемеловых палеориктид возникают две долго конкурировавшие ветви хищных млекопитающих — креодонты и миациды. Первые сейчас рассматриваются в качестве самостоятельного отряда Creodonta, а вторые составляют самое примитивное семейство отряда настоящих хищных (Carnivora). К концу палеогена большинство креодонтов вымирает, и начинается широкая радиация хищных современного типа. Именно с этого времени (около 25 млн. лет назад) известны почти все семейства современных хищных. Вскоре в различных группах хищных осуществляется переход некоторых форм к водному образу жизни. Потомки этих водных хищников,, известные в палеонтологической летописи с миоцена, по традиции объединяются в отряд ластоногих (Pinnipedia), хотя по современным данным это сборная группа (ушастые тюлени и моржи тяготеют к ветви медвежьих, а настоящие тюлени — к куньим).
В конце палеоцена появляются наиболее примитивные представители грызунов (отряд Boden-tia), неуклонно процветавших на протяжении последних 50 млн. лет и Ставших самым обширным отрядом современных млекопитающих (к грызунам относится около 40% всех видов ныне живущих зверей). Настоящие зайцеобразные (отряд Lagomorpha) появляются в палеонтологической летописи несколько позднее, но в палеоцене существовали их вероятные предки, анагалиды; некоторые роды анагалид имеют признаки, которые позволяют исследователям предполагать общее происхождение зайцеобразных и грызунов от одних и тех же палеоценовых предков.
В палеоцене известны вероятные предки современных шерстокрылов — плагиомениды, объединяемые с ними в современный отряд Dermoptera. В последнее время их часто сближают с приматами.
Наиболее примитивные рукокрылые (отряд Chiroptera) известны с начала эоцена; предполагается, что они произошли от насекомоядных отряда Insectivora.
В первой половине палеоцена происходит бурное развитие еще одной ветви плацентарных, связанной с дивергенцией кондиляртр. Большинство сохранившихся до наших дней отрядов этой ветви (китообразные, даманы, хоботные, сирены, парнокопытные, мозоленогие, непарнокопытные), представленные примитивными формами, появляются на протяжении эоцена (37—55 млн. лет назад), но корни их дивергенции уходят„ вероятно, в палеоцен. Так, происхождение примитивных раннеэоценовых китообразных (отряд Cetacea), бывших, возможно, полуводными животными, связывается многими исследователями с наземными палеоценовыми мезонихидами, относимыми сейчас к кондиляртрам. От различных групп кондиляртр, вероятно, произошли непарнокопытные (отряд Perissodactyla), появившиеся в конце палеоцена, и парнокопытные (отряд Artiodactyla), известные с начала эоцена.
Примитивные черты строения трубкозубов (отряд Tubulidentata) также позволяют предполагать их палеоценовое происхождение от кондиляртр, хотя достоверные ископаемые остатки трубкозубов известны лишь с середины миоцена. То же можно сказать и о даманах (отряд Hyracoidea), первые представители которых появляются в палеонтологической летописи в начале эоцена. Одни исследователи сближают этот отряд с кондиляртрами и непарнокопытными, другие — с хоботными и сиренами, но так или иначе это очень древняя группа, сохранившая до наших дней некоторые черты примитивных копытных наряду с приобретенными весьма своеобразными признаками.
Отряды хоботных (Proboscidea) и сирен (Sirenia), также известные с эоцена, сближаются друг с другом по некоторым морфологическим и иммунологическим признакам, но их взаимоотношения с другими отрядами наименее ясны. Наконец, мозоленогие (отряд Tylopoda), которых часто относят к парнокопытным в ранге подотряда, также известны с конца эоцена и благодаря многим своеобразным чертам, приобретенным в ходе длительной эволюции, могут рассматриваться в качестве самостоятельного отряда.
Млекопитающие (лат. Mammalia) — класс позвоночных животных, основной отличительной особенностью которых является вскармливание детёнышей молоком (подрастая, постепенно переходят с молока на твёрдую пищу). Входит в кладу Synapsidomorpha надкласса четвероногих; включает в свой состав и вид Человек разумный (лат. Homo sapiens). В мире известно по разным оценкам от 5000 до 5500 видов млекопитающих (в справочнике «Mammal Species of the World» 2005 года издания говорится о 5416 видах), а на территории России — до 380 видов.
Распространены почти повсеместно. Произошли от продвинутых синапсид в конце триаса. Включают ныне живущих однопроходных, сумчатых и плацентарных, а также большое число вымерших групп. Представления о родственных связях между отдельными отрядами сумчатых и плацентарных претерпели в начале XXI века значительные видоизменения (прежде всего, благодаря достижениям молекулярной филогенетики).
Внешний вид млекопитающих весьма разнообразен, но в целом следует характерному и для других четвероногих плану строения. Анатомия и физиология млекопитающих характеризуется наличием тех же функциональных систем, что и у прочих четвероногих; однако многие из этих систем достигли весьма высокого уровня развития, так что данный класс среди позвоночных является наиболее высокоорганизованным. Весьма гибки и совершенны приспособления млекопитающих к обитанию в различных жизненных средах, сложностью и разнообразием отличается их поведение. Они занимают доминирующие позиции среди наземной фауны (в водной среде они уступают лишь лучепёрым рыбам); велико и многообразно их значение в жизни человека.
Изучением млекопитающих занимается наука териология (маммалиология)/ (Википедия)
Книги:
 Жизнь животных. Том 7. Млекопитающие. / В. Е. Соколов — 1989
Жизнь животных. Том 7. Млекопитающие. / В. Е. Соколов — 1989
 Систематика млекопитающих. Том 1: Однопроходные, Сумчатые, Насекомоядные, Шерстокрылы, Рукокрылые, Приматы, Неполнозубые, Ящеры / В. Е. Соколов — 1973
Систематика млекопитающих. Том 1: Однопроходные, Сумчатые, Насекомоядные, Шерстокрылы, Рукокрылые, Приматы, Неполнозубые, Ящеры / В. Е. Соколов — 1973
 Систематика млекопитающих. Том 2: Зайцеобразные, Грызуны / В. Е. Соколов — 1977
Систематика млекопитающих. Том 2: Зайцеобразные, Грызуны / В. Е. Соколов — 1977
 Систематика млекопитающих. Том 3: Китообразные, Хищные, Ластоногие, Трубкозубые, Хоботные, Даманы, Сирены, Парнокопытные, Мозоленогие, Непарнокопытные / В. Е. Соколов — 1979
Систематика млекопитающих. Том 3: Китообразные, Хищные, Ластоногие, Трубкозубые, Хоботные, Даманы, Сирены, Парнокопытные, Мозоленогие, Непарнокопытные / В. Е. Соколов — 1979
 Жизнь животных. Том 1: Млекопитающие / Брэм А. Э.— 1992
Жизнь животных. Том 1: Млекопитающие / Брэм А. Э.— 1992
 Териология./ Барабаш-Никифоров И. И., Формозов А. Н.— 1963
Териология./ Барабаш-Никифоров И. И., Формозов А. Н.— 1963
 Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989
Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989