



ОПИСАНИЕ:
Отряд Гинкговые / Ginkgoales
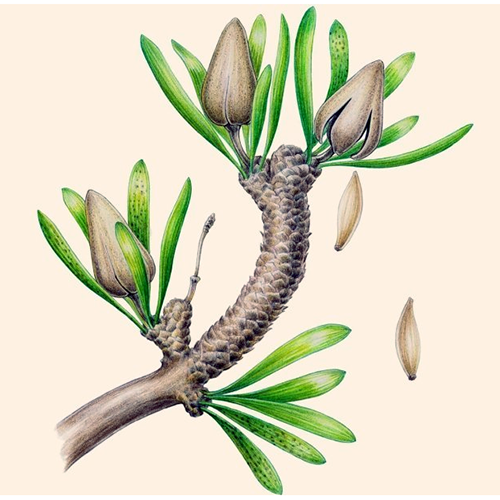
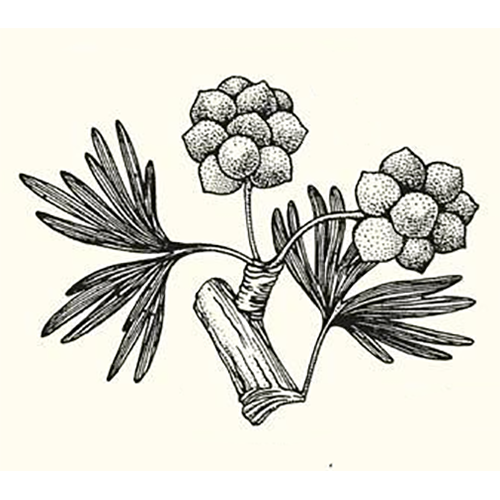
Порядок семенных растений, выделяемый в собственный отдел гинкговидных (Ginkgophyta). В современной флоре представлен единственным видом — двулопастным гинкго (Ginkgo biloba). Наиболее древние находки гинкговых датируются поздним палеозоем.Гинкговые являются двудомными растениями с однополыми репродуктивными органами (стробилами). Микростробил представлен простой осью со спирально прикреплёнными микроспорофиллами, состоящими из тонкой ножки с сидящими на ней пыльцевыми мешочками (микроспорангиями). Внутри пыльцевых мешочков находятся пыльцевые зерна (микроспоры). По своим размерам и форме они похожи на пыльцевые зёрнам беннеттитов и саговников. Женские органы — мегастробилы также имеют простое строение и представляют собой длинную ножку с сидящими на верхушке двумя семязачатками, причем в семя развивается только один из них. Опыление ветром, однако заострённая форма пыльцевых зерен, которые лишены воздушных мешков, плохо способствует этому процессу. При густом стоянии деревьев пыльца, вероятно, стрясывается. Несовершенство опыления рассматривается одной из причин вымирания гинкговых. Процесс слияния мужской половой клетки с женской происходит у гинкговых за счёт подвижных сперматозоидов, что впервые было обнаружено японским художником и ботаником-любителем Хирасэ Сакугоро в 1896 году. Семена овальной формы и длиной 1—3 см. Снаружи они покрыты мясистой оболочкой, окружающей скорлупу из каменистых клеток, под которой находится тонкая плёнка, плотно покрывающая семя. Семена видов, известные в ископаемом состоянии, сходны с семенами современного Ginkgo biloba.
По своему строению основных органов гинкговые наиболее сходны с кордаитовыми. Предполагается, что они произошли от общего предка, а их развитие происходило параллельно. Расцвет гинкговых имел место в юрском периоде. Основной территорией их становления и развития была древняя Ангарида (северо-восточная Азия). На этом материке во время мезозоя гинкговые образовывали обширные лесные массивы и стали основой образования угольных залежей. Ископаемым остатки гинкговых свидетельствуют, что в прошлом эта группа была представлена весьма разнообразно. Гинкговые в целом произрастали преимущественно в умеренном климате, что подтверждается опадающей листвой и наличием годовых колец.
Порядок достоверно известен с триаса, хотя не исключено, что некоторые пермские и даже верхнекарбоновые формы принадлежат к гинкговым.
Единственный современный вид (Ginkgo biloba) представлен лептокаульными пикноксильными листопадными деревьями до 30 м высотой, побеговая система которых сложена ауксибластами и пролептическими брахибластами, способными превращаться друг в друга. Вероятно, что такие же особенности были свойственны и ископаемым гинкговым.
Апекс корня содержит несколько инициален, располагающихся однослойной поперечной пластинкой. Радиальный пучок корня с диархной ксилемой. Апекс побега также содержит несколько инициален, в расположении которых слоистость не выражена. В стеблях развита эвстела с эндархной протоксилемой в коллатеральных пучках. Листовые следы двухпучковые, причем пучки одного листового следа ответвляются от смежных стеблевых пучков. В осевых органах протоксилема состоит из спиральных трахеид с примесью кольчатых, а метаксилема — из точечных трахеид. Вторичная ксилема сложена точечными трахеидами с однорядными округлыми окаймленными порами на радиальных стенках в ранней древесине и на тангенциальных стенках — в поздней. Лучи гомогенные однорядные малослойные. Поры на полях перекреста значительно мельче межтрахеидальных пор и имеют эллиптические, а не круглые отверстия. Замыкающая пленка некоторых межтрахеидальных пор утолщена в центре в небольшой торус. Вторичная флоэма содержит ситовидные клетки с поперечно вытянутыми ситовидными полями, тяжевую паренхиму с кристаллоносными идиобластами, волокна и гомогенные однорядные малослойные лучи. На периферии непроводящей зоны из клеток тяжевой паренхимы развиваются многочисленные склереиды.
Трофофиллы современного гингко простые, веерные, с открытым дихотомическим жилкованием листовой пластинки. Форма и степень расчленения листовой пластинки варьируют с возрастом побега и всего растения и несколько различаются на аукси- и брихибластах. У ископаемых представителей листовые пластинки были в целом более сильно расчлененными. Протоксилема в проводящих пучках жилок листа мезархная. Наряду с трофофиллами есть катафиллы, представленные почечными чешуями.
В кортексе, основной паренхиме черешков и в мезофилле находятся слизевые вместилища.
Гинкго — двудомное анемофильное растение. Одни особи продуцируют безмешковую пыльцу, а другие — семена. «Пол» спорофита определяется посредством половых хромосом. Мужские фруктификации развиваются на брахибластах в пазухах нескольких катафиллов. Каждая фруктификация состоит из удлиненной повислой оси, на которой по неправильной спирали располагаются пельтатные микроспорофиллы с сильно редуцированным щитком и обычно с двумя висячими микроспорангиями (ископаемые Ginkgoales имели менее редуцированные щитки, несшие соответственно большее число спорангиев). Вскрывание микроспорангиев обеспечивают клетки 2—3 слоев субэпидермального эндотеция, имеющие спиральные утолщения стенок. Их гигроскопическая деформация вызывает разрыв стенки микроспорангия продольной щелью. Пыльцевые зерна эллипсоидальные, лодочковидные, моносулькатные, причем ширина сулькуса значительно различается в зависимости от степени гидратированности зерна.
Женские фруктификации также развиваются на брахибластах в пазухах катафиллов. Каждая из них в типе состоит из довольно длинной оси с двумя атропными семяпочками на верхушке. По оси проходят четыре проводящих пучка, по два к каждой семяпочке. Иногда у современного гинкго, как и у ископаемых гинкговых, бывает ветвистая женская фруктификация с несколькими семяпочками, к каждой из которых подходит пара проводящих пучков. Интегумент закладывается двумя при- мордиями с противоположных сторон от нуцеллуса и затем долго сохраняет «двурогость». В зрелой семяпочке интегумент слит с нуцел- лусом более чем наполовину, а на вершине имеет узкий микропилярный канал. В интегументе проходят два пучка, продолжающиеся из оси фруктификации. В основании семяпочки имеется небольшой валик, гомология которого неясна. Судя по некоторым уродливым фруктификациям, он может быть остатком пластинки мегаспорофилла, точнее, пельтоида или капсулы предполагаемых предковых форм.
К моменту опыления, весной, из верхушки нуцеллуса путем частичного разрушения клеток формируется короткий сальпинкс. К этому времени в нуцеллусе только завершается мегаспорогенез, и образуется линейная тетрада мегаспор. Сальпинкс выделяет опылительную каплю, выступающую из микропиле и улавливающую разносимые ветром пыльцевые зерна. Позднее из-за подсыхания капли и поглощения ее клетками нуцеллуса она втягивается вместе с попавшими пыльцевыми зернами внутрь семяпочки, после чего микропиле зарастает, а семяпочка начинает развиваться в семя. Почти всегда в каждой паре семяпочек развивается только одна.
После опыления, но задолго до оплодотворения интегумент преобразуется в тесту. В нем дифференцируются три зоны. Наружная экзотеста из сочной паренхимы (саркотеста), вероятно, предназначена для привлечения животных1, проглатывающих семена и таким образом эндозоохорно распространяющих непереваренные зародыши. Однако в современной фауне неизвестны животные, распространяющие семена гинкго. Возможно, что они вымерли. Средняя зона — мезотеста — выполнена склеренхимой (склеротеста), защищающей зародыш от переваривания в кишечном тракте животных. Внутренняя тонкая эндотеста представлена паренхимой (пахитеста), ее функции непонятны.
Развитие женского гаметофита начинается после опадения семени и занимает около двух месяцев. При этом из халазальной мегаспоры вырастает це- ноцит, постепенно разрушающий и замещающий нуцеллус. В ходе'кариоки- незов в ценоците образуется более 1000 ядер, после чего начинается центрипетальная целлюляризация и возникает многоклеточный паренхимный гаметофит. Он имеет зеленую окраску, обусловленную наличием в клетках хорошо развитых хлоропластов. На микропилярном конце гаметофита образуются обычно два архегония. Каждый архегоний развивается из одной поверхностной клетки, делящейся периклинально. Из наружной клетки образуются две клетки шеики, а внутренняя клетка начинает расти и делится неравно на мелкую брюшную канальцевую клетку и яйцеклетку; шейковые канальцевые клетки не образуются.
Первые стадии онтогенеза мужского гаметофита проходят эндоспорически в микроспорангии. Микроспора делится резко неравно с образованием с проксимальной стороны мелкой первой проталлиальной клетки, вскоре отмирающей. Оставшаяся крупная клетка также делится неравно на мелкую вторую проталлиальную клетку, прилежащую к остаткам первой, и крупную антеридиальную инициале. Последняя опять делится неравно на более крупную сифоногенную (гаусториальную, или вегетативную) клетку, расположенную дистально, и мелкую антеридиальную клетку. Из всех этих клеток мужского гаметофита только проталлиальные возможно достаточно уверенно гомологизировать с вегетативными клетками менее редуцированных гаметофитов растений других таксонов. Точные гомологии остальных клеток пока не выяснены. Хотя в их названии фигурирует апелляция к антеридию, но неизвестно, есть ли на самом деле у Ginkgo какие бы то ни было гомологи антеридия. После образования антеридиальной и сифоногенной клеток в развитии мужского гаметофита наступает пауза, и он высеивается из микроспорангия.
Только гаметофиты, попавшие внутрь семяпочки, возобновляют развитие. После опыления верхушка нуцеллуса постепенно разрушается с образованием заполненной жидкостью пыльцевой камеры, которая, углубляясь, через 2,5 — 3 месяца достигает женского гаметофита, микропилярная поверхность которого становится дном камеры. К этому моменту, т.е. в уже отделившемся от материнского растения семени, женский гаметофит развивает архегонии, а мужской гаметофит начинает прорастать пыльцевой трубкой. Пыльцевая трубка растет, ветвясь, по поверхности и частично по паренхиме нуцеллуса в сторону от пыльцевой камеры. Она выполняет гаусториальную функцию, усиленно поглощая низкомолекулярные углеводы. После прорастания пыльцевого зерна антеридиальная клетка делится на прилежащую к проталлиальной клетке клетку-ножку и обращенную к сифоногенной клетке генеративную клетку. Еще примерно через месяц последняя делится на два сперматозоида кубаревидной формы со спиралью многочисленных жгутиков на переднем конце.
После завершения сперматогенеза пыльцевая трубка в основании, где располагаются сперматозоиды, лопается, изливая содержимое в пыльцевую камеру. Это приводит к заметному повышению концентрации сахаров в жидкости, заполняющей камеру, раствор которых оказывается гипертоническим по отношению к клеточному соку клеток шеек архегониев. Вследствие этого клетки шейки теряют тургор и раскрывают канал шейки. Сперматозоиды в жидкости пыльцевой камеры активно плывут к архегониям, но лишь в одном из них происходит половой процесс. Из зиготы через стадию ценоцита с примерно 256 ядрами развивается зародыш с двумя семядолями. Ткань женского гаметофита в виде (первичного) эндосперма сохраняется и в зрелом семени, занимая большую часть его объема. Семена не имеют периода глубокого покоя, что принято считать признаком примитивности. При прорастании семядоли остаются внутри семени (гипогейное прорастание) и выполняют гаусториальную функцию. Первые несколько листьев проростка — чешуевидные катафиллы, и лишь затем формируются характерные вееровидные листья.
Ginkgo biloba — одно из древнейших культивируемых растений Китая и Японии, в течение нескольких тысячелетий разводимое около храмов. Естественный ареал гинкго точно не установлен, но, вероятнее всего, он ограничен участками влажного полувечнозеленого среднегорного леса в Сычуани. В настоящее время гинкго широко культивируют как парковое и аллейное дерево в областях умеренного теплого климата. Кроме того, его листья и семена используют в медицине.
Гинкговые, по-видимому, произошли от мезозойских Peltaspermatales, имевших пельтоиды как на мужской, так и на женской фруктификациях. В эволюции гинкговых произошло сильное упрощение листьев и фруктификаций с почти полной редукцией пельтоидов.
Книги:
 Ботаника, Том 3, Высшие растения, Тимонин А.К., 2007
Ботаника, Том 3, Высшие растения, Тимонин А.К., 2007
 Ботаника, Том 4, Книга 1, Систематика высших растений, Тимонин А.К., Филин В.Р., 2009
Ботаника, Том 4, Книга 1, Систематика высших растений, Тимонин А.К., Филин В.Р., 2009
 Ботаника, Том 4, Книга 2, Систематика высших растений, Тимонин А.К., Соколов Д.Д., Шипунов А.Б., 2009
Ботаника, Том 4, Книга 2, Систематика высших растений, Тимонин А.К., Соколов Д.Д., Шипунов А.Б., 2009
 Фёдоров А.А. Жизнь растений в шести томах. Том 4. Мхи. Плауны. Хвощи. Папоротники. Голосеменные растения
Фёдоров А.А. Жизнь растений в шести томах. Том 4. Мхи. Плауны. Хвощи. Папоротники. Голосеменные растения
 Забинкова Н. Н., Кирпичников М. Э. Латинско-русский словарь для ботаников. — М., Л.: Изд-во АН СССР, 1957. — 335 с.
Забинкова Н. Н., Кирпичников М. Э. Латинско-русский словарь для ботаников. — М., Л.: Изд-во АН СССР, 1957. — 335 с.
 Фёдоров Ал. А., Кирпичников М. Э. и Артюшенко З. Т. Атлас по описательной морфологии высших растений. Стебель и корень
Фёдоров Ал. А., Кирпичников М. Э. и Артюшенко З. Т. Атлас по описательной морфологии высших растений. Стебель и корень
 Фёдоров Ал. А., Кирпичников М. Э. и Артюшенко З. Т. Атлас по описательной морфологии высших растений. Лист
Фёдоров Ал. А., Кирпичников М. Э. и Артюшенко З. Т. Атлас по описательной морфологии высших растений. Лист