

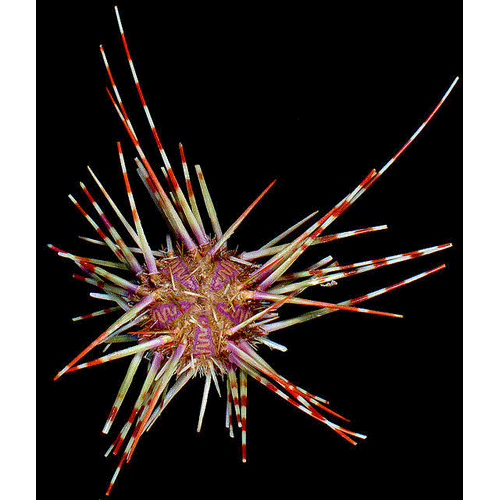


Описание:
Иглокожие представляют собой самостоятельный и весьма своеобразный тип животного мира. По плану строения они совершенно не сравнимы ни с какими иными животными и благодаря особенностям организации и оригинальной форме тела, напоминающей звезду, цветок, шар, огурец и пр., издавна привлекали к себе внимание. Название «иглокожие» дано еще древними греками. В античное время уже различали среди животного мира морских звезд, голотурий и морских ежей, и история изучения этих животных связана с именами Аристотелями Плиния. В XVIII и в начале XIX в. ими занимались такие видные ученые, как Клейн, Линней, Ламарк и Кювье. Однако большинство старых зоологов относили иглокожих либо к червям, либо к кишечнополостным.Название лучистые, или зоофиты, сохраняется за иглокожими вплоть до 1847 г., когда Ф р е й и Л е й к а р т выделили их в самостоятельный тип, указав на значительные отличия иглокожих от лучистых, в частности от кишечнополостных животных.
Последующие работы таких ученых, как Л ю д-в и г, А г а с с и ц, А. О. К о в а л е в с к и й, И. И. Мечник о в, Тиль, Безер, Фишер, Мортенсени др., значительно расширили представление о систематике, анатомии и эмбриологии иглокожих, о родственных отношениях между отдельными представителями этого типа. И. И. Мечников, изучая развитие иглокожих, установил их родство с кишечножаберными и доказал, что ветвь иглокожих близка ветви хордовых животных. Датским ученым Мортенсеном была не только разработана система для современных и ископаемых классов иглокожих, но п дана классификация их личинок параллельно системе взрослых. Это особенно важно, так как личинки иглокожих очень своеобразны, весьма отличны от взрослых форм, ведут пелагический образ жизни и долгое время считались самостоятельными организмами.
Среди советских исследователей, особенно много сделавших для изучения иглокожих, прежде всего необходимо отметить Л. М. Д ь я к о и о в а и Д. М. Ф е д о т о в а.
За последние 20 лет значительно активизировали свои исследования палеонтологи. В американском многотомном издании «Палеонтология беспозвоночных», большое внимание уделено и иглокожим. Было обнаружено много новых форм. Все это привело к пересмотру системы иглокожих.
С начала XX в. (со времени Б е з е р а) в науке прочно установилось деление иглокожих на два подтипа: стебельчатые иглокожие (Pelraatozoa) и свободноживущие иглокожие (Eleuterozoa). В типе насчитывали 13 классов. Такое деление и сейчас находит много сторонников, особенно среди эмбриологов и зоологов. После работ палеонтологов, среди которых надо прежде всего отметить работы Моора, Фелла, Убагса, Дарема, Филипаи др., все больше сторонников завоевывает система деления иглокожих на 4 подтипа: гомалозои (Homalozoa), пелъматозои (Pelmatozoa), эхинозои (Echinozoa) и астерозои (Asterozoa), включающих более 20 классов.
Иглокожие появились на Земле давно, свыше 580 млн. лет назад. Обладание известковым скелетом способствовало довольно хорошему сохранению их в ископаемом состоянии. Остатки этих животных находят уже в отложениях раннего кембрия. Древние иглокожие достигали в палеозойскую эру значительного разнообразия форм. Здесь имелись представители 22 классов всех 4 подтипов. Наиболее древние и примитивные классы относились к подтипу гомалозой: ктеноцистоидеи (Ctenocystoidea), гомостелии (Homostelea), стилофоры (Stylophora) и гомоиостелии (Homoios-telea). Они имели мешковидное сдавленное тело, лишенное радиальной симметрии, и обладали рядом признаков, сближающих их с низшими хордовыми. Гомалозои сильно обособлены от остальных иглокожих, они появились в кембрии и исчезли в девоне. Характерные же черты типа иглокожих получают развитие у представителей подтипов пельматозой, эхинозой и астерозои.
Среди 7 классов пельматозой наиболее древними были эокриноидеи (Hocrinoidea), известные с кембрия до силура (риг. 11Г>). От них, вероятно, произошли известные с ордовика классы ромбифер (Hombifera), днплопорит (l)iploporita)1, бластоидей (Blast oidea), иарабластоидей (Parablasto-idca) и паракринопдей (Paracrinoidea). В связи с прикреплением к субстрату у них развилась радиальная симметрия, причем можно было наблюдать появление пятилучевой симметрии. Они господствовали в морях 500—400 млн. лет назад, ромбиферы и дпплопориты исчезли в девоне, а бластоидеи — в перми.
Среди всех пельматозой только виды класса морских лилий (Crinoidea) встречаются в современной фауне. Вымершие морские лилии, представленные в ордовике, силуре, девоне и карбоне (согласно Фишеру. 5 тыс. видов), почти полностью вымерли к концу палеозоя. Однако в начале мезозоя, приблизительно 200—180 млн. лет назад, появилась новая группа морских лилий — подкласс артикулят (Arliculala), представители которого не только сохранились до наших дней, но и достигают большого расцвета в настоящее время.
Эволюция древних классов подтипа эхинозой тоже прошла в палеозое, и до нашего времени дошло лишь 2 класса из 8, отнесенных к этому подтипу. Среди вымерших классов эхниозой наиболее древними были раннекембрииские камптостроматоидеи (Caniplostromaloidea) и геликоплакоидеи (Ilelicoplacoidea). Последние имели веретенивидный, сжимавшийся и расширявшийся панцирь со спирально расположенными складками, рот и апикальное поле, помещавшиеся на противоположных полюсах.
Существовавшие с кембрия до перми эдриоастероидеи (Ivlrionsleioidea) уже обладали пятилучевой симметрией. Они располагались на субстрате ртом кверху. Большинство из них прирастало к субстрату широкой поверхностью тела, и только очень немногие имели стебелек или вообще не были прикреплены. Ранее их включали в подтип пельматозой и считали возможными предками морских звезд. Последнее время палеонтологи, учитывая предположения Фелла, выводят их пз общего зхинозойпого ствола. От них, возможно, произошли эдриобластопдеи (Rdrioblastoidca) и никлоцистоидеи(Cyclocysloidea), известные из ордовика.
Офиоцистоидеи (Ophiocistioidea), которых найдено около 10 видов, появились в ордовике и в карбоне исчезли. Тело их имело дисковидную или шаровидную форму, было лишено выступающих лучей и сплошь покрыто пластинками. Рот помешался в центре стропы, обращенной к субстрату, и был снабжен крепкими подвижными челюстями. Происхождение офноцнстиоидей не ясно, но они обнаруживают некоторое сходство с морскими ежами.
Из всех классов эхииозой только морские ежи (Echinoidea) и голотурии (Holotluirioidea), возникшие в ордовике свыше 450 млн. лет назад, встречаются и в современной фауне. Самыми древними морскими ежами были представители небольшого отряда ботриоцидароид (Bolhrioci-daroida), которые вымерли в верхнем ордовике, не оставив потомства. Наиболее многочислен пой среди палеозойских морских ежей была группа отряда лепидоцентроид (Lepidocentroida), достигшая расцвета в карбоне и вымершая в перми. С лепидоцентроидами очень сходны современные ежи семейства эхпиотурпид, правда, найденные в ископаемом состоянии только с юры. Почти все остальные современные ежи начинают эволюцию тоже с юры, но виды отряда цидароид (Gydaroida) появились значительно раньше, уже в девоне. Расцвет их приходится на мезозой, а в современной фауне они представлены относительно небольшим числом видов.
Геологическая история голотурпй известна мало вследствие отсутствия у них сплошного скелета. Однако это древняя группа, так как спикулы (склериты) вымерших голотурпй известны из среднего ордовика. Вполне возможно, что это были животные с удлиненной формой тела, с кожистыми покровами, без сплошного наружного скелета.
Последпий подтип — астерозои — представляет собой наиболее компактную группу. Происхождение его остается неясным, но все виды класса подтипа — сомастеропдеи (Somasleroidea), морские звезды (Asleroidea) и офиуры (Opliinroidea) — известны с ордовика и имеют много общего. Все опи звездообразной формы, и многие видят в сомастероидеях (с их легким, подвижным скелетом) предков морских звезд и офнур. Вымершие морские звезды имели грубый, прочный скелет, состоявший из отдельных пластинок. В ископаемом состоянии обнаружено около 500 видов. В конце мезозоя отмечен некоторый подъем в их развитии, и сейчас это процветающий класс.
Древние офиуры обладали рядом признаков, сближающих их с морскими звездами. Они имели открытую амбулакральную борозду и не имели характерных черт настоящих офиур. Среди них встречались виды, которые были гигантами даже по сравнению с позднейшими типичными офиурами. Такие офиуроподобные формы вымерли в верхнем карбоне, где возрастает число видов настоящих офиур, по заметная их эволюция начинается в мезозое и продолжается вплоть до настоящего времени. В современных морях офиуры наиболее многочисленны по числу видов среди всех иглокожих.
Вымерших иглокожих обнаружено свыше 13 ООО видов, а ныне живущих насчитывается немногим более 1000 видов.
Среди современных иглокожих различают 5 классов:
Размеры тела иглокожих обычно колеблются в пределах от 5 до 50 см, но имеются виды, длина которых не превышает нескольких миллиметров, а у других, наоборот, может доходить до 5 м.
Известные нам классы иглокожих очень своеобразны и весьма отличаются друг от друга, но, несмотря на большое многообразие форм, они имеют целый ряд признаков, свойственных только этим животным, которые указывают как на общность происхождения всей группы, так и на своеобразный путь ее эволюции. Действительно, лишь иглокожие обладают особой водоносной (амбулакральной) системой, состоящей из серии тонкостенных каналов, наполненных жидкостью, имеют лучистый план строения, обычно кратный 5, и сильно развитый скелет, который по происхождению, в отличие от скелетов других беспозвоночных животных, является внутренним.
Амбулакральная с и с т е м а у большинства форм служит для движения, осязания, а у морских лилий, офиур и некоторых ежей выполняет дыхательную функцию. Она в наиболее типичном случае состоит ил к о л ь ц е в о г о к а н ал а, окружающего ротовое отверстие с внутренней стороны, и 5 р а д и а л ь и ы х к а н а л о в, отходящих от кольцевого и заканчивающихся слепо или небольшим чувствительным отростком. Радиальные каналы, проходя по внутренней стороне луча, дают попарно боковые веточки к ножкам, которые называются а м б у л а к р а л ь н ы м п. Амбулакральные ножки представляют собой тонкие цилиндрические, очень растяжимые трубочки, обычно снабженные па конце присоской, реже лишенные ее. У основания каждой ножки находится тонкостенный пузырек, или а м п у л а. Между радиальными каналами (интеррадиально) от кольцевого канала отходят тонкостенные, сильно растяжимые, часто очень больших размеров мешки — и о л и е-в ы и у з ы р и, служащие резервуарами жидкости амбулакральиой системы. Число полпевых пузырей сильно варьирует, иногда их бывает несколько даже в одном ннтеррадиальном промежутке, у никоторых форм они вообще могут отсутствовать. Межрадиально с кольцевым каналом соединяется непарный к а м е н и с т ы й к а н а л, названный так за твердость своих обылвествленных стенок. Каменистый канал соединяется с лежащей у большинства форм на поверхности тела особой м а д р е п о р о в о п п л а с т п и к о й или м а д р е п о р и т о м. Мадрепоровая пластинка пронизана мельчайшими отверстиями -— порам и, через которые в амбулакральную систему поступает морская вода.
Детали строения и функции амбулакральной системы могут претерпевать ряд изменений как у различных классов, так и в пределах одного класса, отряда и даже семейства иглокожих. Так, у части голотурий радиальные каналы укорачиваются до коротких обрубков или вообще исчезают. У некоторых морских звезд, офиур и голотурий бывает несколько каменистых каналов и несколько мадрепоровых пластинок, причем у большинства голотурий каменистый канал теряет сообщение с наружной средой и вместе с мадрепоритом свисает в полость тела. У морских звезд, ежей и представителей одного отряда офиур кольцевой канал имеет особые гроздевидные или мешковидные придатки, состоящие из слепых простых пли разветвленных трубочек. Это т и д е м а н о вы те л ь ц а, назначение которых точно не установлено, но предполагают, что в них образуются целомоциты, играющие важную роль в очищении целомической жидкости и амбулакральной системы. Особенно большие изменения в строении и функции претерпевают амбулакральные ножки. Например, у морских лилий они лишены ампул, присасывательных дисков, остроконечны, никогда не служат органами движения, а выполняют чувствительную и дыхательную функции. Две пары ножек, расположенные около рта, превращаются в ротовые щупальца, помогающие при захвате пищи. У многих голотурий у одного и того же экземпляра встречаются как двигательные ножки, снабженные присосками, так и ножки, лишенные присосок, превращенные в сосочки и выполняющие уже другие функции, а у представителей двух отрядов голотурий ножки редуцируются. Ротовые щупальца голотурий представляют собой видоизмененные ножки. Среди морских ежей встречаются виды с разнообразными по форме и функции амбулакральными ножками. У них бывают наряду с двигательными ножками конические, розеткообразные, кистевидные ножки, которые несут уже иные функции.
Несмотря на такое изменение ножек, основное их значение у морских звезд, большинства голотурий и ежей — передвижение по дну морей. В этом случае жидкость амбулакральной системы гонится из кольцевого канала в радиальные, откуда по боковым ветвям поступает в ампулы ножек. Благодаря сокращению мышц жидкость из ампул перегоняется в ножки, которые от этого сильно вытягиваются, и подошвы ножек соприкасаются с субстратом. От этого соприкосновения сокращаются мышцы подошвы (присоски), последняя вдавливается, и между подошвой и субстратом образуется разреженное пространство, обусловливающее прикрепление ножки к субстрату. Прикрепление ножек к субстрату благодаря давлению воздуха и воды и действию клейкой слизи, выделяемой железистыми клетками присоски, столь прочно, что часто при попытке оторвать прикрепившееся животное его ножки обрываются и остаются связанными с субстратом. Последующее сокращение прикрепившихся ножек подтягивает тело животного, а жидкость из сократившейся ножки вновь поступает в расслабленную к этому времени ампулу. Клапаны, находящиеся в ампулах, регулируют наполнение их жидкостью.
Другой характерной чертой иглокожих является л у ч е в а я с и м м е т р и я их тела. Число лучей обычно кратно 5. Тело иглокожих поделено на 10 частей (секторов). Сектор ы, в которых расположены амбулакральные ножки и радиальные каналы амбулакральной системы, называются а м б у л а к р а м и или радиусами. Они чередуются с м е ж р а д и а л ь н ы м и сектора м и, которые называются интеррадиусами или и п т е р а м б у л а к р а м и. У разных классов иглокожих радиусы и интеррадиусы выражены различно. У одних они слиты с общим телом животного (голотурии и морские ежи), у других радиусы могут сильно вытягиваться, образуя подвижные или неподвижные лучи, иногда называемые руками. В таких случаях животные приобретают звездообразную форму (звезды, офиуры). Иногда лучи древовидно разветвляются, как у ветвящихся офиур, или становятся перистыми, как у морских лилий. У звездообразных форм число лучей тоже может меняться от 5 до 10 и даже до 50. Изредка среди обычных пятилучевых форм могут попадаться четырех- и шестилучевые экземпляры.
Не менее характерной чертой типа иглокожих, наряду с амбулакральной системой и лучистым планом строения, является наличие у них известкового скелета, заложенного в соединительнотканном слое кожи. Кожа иглокожих состоит из однослойного покровного эпителия, толстого слоя кутиса, или собственно кожи, и слоя ресничного эпителия, ограничивающего вторичную полость тела, или ц е-л о м. Наружный покровный эпителий у большинства форм (исключение составляют голотурии) снабжен ресничками, расположенными неравномерно. Действие этих ресничек вызывает возникновение на поверхности тела животного токов воды, идущих в определенных направлениях и служащих для подачи пищи, для дыхания и очищения тела от грязи. В наружном эпителии кожи имеется большое количество железистых клеток, выделяющих слизистый, клейкий или ядовитый секрет, а у некоторых видов встречаются железистые клетки, секрет которых вызывает свечеиие организмов. Здесь также находятся скопления пигментных клеток, придающих иглокожим удивительно красивую и яркую окраску. Кутис, подстилающий наружный эпителий, состоит из соединительной ткани, в которой можно различить три слоя. В нижнем, внутреннем, слое кутиса располагаются мышцы, сильно развитые у голотурий, у которых они входят в состав кожномускульного мешка, и значительно редуцированные у морских лилий, ежей и большинства офиур в связи с развитием у них прочного скелета. В наружном слое кутиса, под эпителием, образуются скелетные элементы, поэтому по своему происхождению из мезодермального слоя они скорее соответствуют покровным костям позвоночных животных, чем раковине улиток.
Для построения скелета иглокожие используют карбонат кальция, растворенный в морской воде. По происхождению скелет всех иглокожих является внутренним и возникает в виде мелкого, едва заметного под микроскопом зернышка карбоната кальция внутри клеток соединительной ткани. Это зернышко растет,- приобретает трехлучевую форму, выпадает ил давшей ему начало клетки и становится межклеточным известковым тельцем. При дальнейшем развитии лучи известкового тельца начинают ветвиться, соседние ветви соединяются друг с другом, замыкая отверстия и образуя мелкие продырявленные пластиночки. Более крупные пластинки скелета формируются путем слияния краев многих мелких пластиночек, поэтому они всегда имеют губчатую структуру. Форма образующихся скелетных элементов специфична и передается по наследству. Она зависит от той роли, которую скелетные элементы выполняют в организме. Хотя форма скелетных элементов иглокожих весьма разнообразна, но всем им присущи оптические и кристаллографические свойства одного кристалла известкового шпата. Поэтому, подобно последнему, каждый скелетный элемент иглокожего обладает двояким лучепреломлением. Однако скелетные элементы иглокожих не являются просто кристаллами известкового шпата, так как у них плотность и показатель преломления меньше, чем у кристаллов известкового шпата. Это биокристаллы, которые возникают в плазме клеток, главным образом из кальцита, с незначительным добавлением органической массы. Поэтому биокрпсталлы не ограничены плоскостями кристалла, а форма их определяется деятельностью внутриклеточной образовательной плазмы.
Известковый скелет иглокожих может быть развит слабо и состоять из отдельных очень маленьких пластинок чрезвычайно разнообразной формы, что и наблюдается почти у всех голотурий, а также в амбулакральных ножках морских ежей, либо достигать мощного развития (все остальные классы иглокожих). В последнем случае он состоит из хорошо заметных пластинок, которые могут скрепляться неподвижно друг с другом наподобие плотной скорлупы, либо образовывать более или менее рыхлую или плотную сеть, либо соединяться друг с другом подобно позвонкам. Пластинки скелета довольно часто несут на поверхности разнообразные иглы (отсюда и название типа), бугры, мелкие гранулы или особые хватательные органы — п е д и ц е л л я р и и.
Среди скелетных элементов, пожалуй, самыми примечательными являются педицеллярпи. Они встречаются у морских звезд и морских ежей иногда в очень большом количестве и представляют собой видоизмененные иглы. Часто педицеллярии достигают такой сложности, что трудно бывает себе представить их происхождение от самой обычной иглы скелетной пластинки. Так, у морских звезд, наряду с самыми простыми педицелляриями, сидящими непосредственно на теле животного и представляющими собой 2 иглы, наклоненные друг к другу, либо две ( д в у с т в о р ч а-тые п е д и ц е л л яри и) или несколько створок, часто расположенных в ряд (гребенчатые педицеллярии), встречаются педицеллярпи с более удлиненными, очень подвижными створками, снабженными зубцами, выростами и сидящими на особом основании. Такие педицеллярии часто образуют пучки или валики вокруг игл и могут прикрепляться к телу животного при помощи мягкой шейки или стерженька. Они очень подвижны, снабжены мышцами, захлопывающими и раскрывающими створки. Основная функция таких педицеллярий — очистка кожных покровов от посторонних частиц, защита мягких частей звезд от повреждений различными мелкими животными. Часто педицеллярии служат и для захвата добычи. У морских ежей педицеллярия верхней стороны панциря, схватив мелкое животное, передает его соседней педицеллярии, та — следующей, и так по всему панцирю вплоть до рта. Иногда педицеллярии используются как органы нападения или защиты и тогда бывают снабжены ядовитой железой. Ядовитые педицеллярии встречаются у морских ежей вместе с другими типами педицеллярий, получившими дальнейшее развитие и усложнение организации по сравнению с педицелляриями морских звезд.
Скелетные пластинки иглокожих очень часто имеют лучевое расположение. Радиальное строение у иглокожих проявляется не только во внешних формах или в размещении пластинок скелета, по и в расположении внутренних органов. Так, нервная и кровеносная системы иглокожих, подобно рассмотренной выше амбулакральной системе, построены по радиальному типу и имеют сходное с ней расположение.
Нервная система иглокожих довольно примитивна, состоит из трех отделов, два из которых, называемые гипоневральной и апикальной системами, являются преимущественно двигательными. Они развиты весьма различно и подчас очень слабо в разных классах данного типа животных, тогда как третий, чувствительный отдел (эктоневральная система) постоянен и хорошо развит у всех иглокожих. Каждый из названных отделов нервной системы состоит из нервного кольца и отходящих от него радиальных нервных стволов, число которых соответствует числу амбулакров. Кольцо и нервные стволы состоят из нервных клеток и нервных волокон и представляют собой нервные центры, воспринимающие и проводящие раздражения. Наиболее поверхностно на ротовой стороне животного расположен чувствительный (эктоневральный) отдел нервной системы, залегающий у морских лилий и морских звезд непосредственно в наружном эпителии амбулакральных желобков, а у офиур, морских ежей и голотурий — в эпиневральных каналах, образованных погружением стенок амбулакральных желобков внутрь и замыканием их в трубки. Нервное кольцо эктоневральной системы окружает глотку пли пищевод, а радиальные стволы у большинства форм доходят до конца амбулак-ров, повторяя сложные разветвления лучей и посылая веточки к различным органам. Глубже зктоневральной системы расположен второй, двигательный отдел нервной системы — гипоневральная система. Между эктоневральной и гипоневральной системами находится тонкий слой соединительной ткани. У голотурий эктоневральная и гипоневральная системы так тесно примыкают друг к другу, что долгое время их даже не различали. Наиболее значительно гипоневральный отдел нервной системы развит у морских лилии и офиур в связи с усиленной подвижностью лучей, причем у офиур радиальные нервы образуют ганглиозные утолщения, посылающие нервы к мышцам позвонков лучей. Третий отдел — апикальная система — расположен на стороне, противоположной ротовой, и связан с зпителием полости тела. Апикальная система отсутствует у голотурий, слабо развита у морских звезд, морских ежей, офиур и только у морских лилий получила достаточное развитие. У морских лилий она ведает движением лучей.
Благодаря такому строению нервной системы, когда преобладающее развитие получает эктоневральный отдел, многие иглокожие становятся весьма чувствительными к внешним раздражениям, хотя органы чувств у них в целом развиты слабо. Многочисленные чувствительные клетки, одиночные или собранные в группы, имеющиеся на амбулакральных ножках морских звезд, офнур, морских ежей и голотурий, па коже рук морских лилий, на ротовых щупальцах и т. д., служат органами осязания. Специальные органы зрении встречаются сравнительно у немногих иглокожих, хотя все эти животные обладают значительной светочувствительностью. Наиболее развитые глаза, построенные по типу глазных ямок, имеются у большинства морских звезд. Они расположены на конце каждого луча у основания концевого щупальца радиального канала водоносной системы. Более слабо развитые глаза встречаются в передней части тела некоторых голотурий. Такие глаза определяют лишь степень яркости, но не различают отдельных предметов. Пигментные пятна морских ежей, расположенные, подобно глазкам морских звезд, над концами радиальных каналов водоносной системы, тоже весьма чувствительны к свету и могут рассматриваться как редуцированные глаза. Несмотря на то что световое раздражение проводится главным образом через глаза, удаление их не лишает животное светочувствительности. Так, весьма обычная у нас в Белом море морская звезда Asterias nibens с отрезанными вместе с глазками кончиками лучей продолжает уползать от очень яркого света и очень сильной темноты. Да и остальные иглокожие, например офиуры, у которых нет органов зрения, тем не менее реагируют на изменение освещённостн, так как вся кожа животных этого типа обладает диффузной восприимчивостью к свету.
Чувства обоняния и вкуса у иглокожих развиты хорошо. Они могут воспринимать вкусовые раздражения даже на значительных расстояниях, хотя органы обоняния и вкуса у них развиты слабо и представлены многочисленными чувствительными клетками, расположенными на амбулакральных ножках, щупальцах офиур и голотурий, ротовых ножках некоторых морских ежей. Некоторые зоологи считают органами вкуса также небольшие, едва заметные глазу образования — сферидии, встречающиеся почти у всех морских ежей. Однако в последнее время большинство исследователей принимают их за органы равновесия. Они обычно расположены близко ко рту на середине амбулакров и состоят из плотного, сильно преломляющего свет вещества, покрытого ресничным эпителием. Сферидии соединены с панцирем коротким стебельком, в основании которого находятся чувствительные клетки и сплетения нервов.
Статическими органами, позволяющими животному определять, где верх и где низ, являются оюцисты, встречающиеся у представителей двух семейств голотурий, которые живут на больших глубинах или зарываются в грунт. Отонисты имеют форму пузырька, содержащего несколько или много мелких отолитов, и связаны непосредственно с радиальными нервами.
Кровеносная система иглокожих также построена но лучевому типу. Она состоит из околоротового кольца и пяти радиальных кровеносных сосудов, идущих в амбулакры между радиальными нервными стволами и каналами водоносной системы. У морских звезд, морских ежей и офиур па стороне, противоположной ротовой имеется второе кольцо кровеносной системы, которое дает боковые ветви к половым железам и соединяется с первым кольцом особым осевым органом, характерным лишь для иглокожих животных. Строение его будет рассмотрено дальше. Кровеносная система у большинства иглокожих представляет собой систему скорее лакун, чем кровеносныx сосудов, и образована просветами в рыхлой соединительной ткани. Жидкость кровеносной системы по составу близка к жидкости полости ала и амбулакральной системы. Основное назначение кровеносной системы иглокожих — разносить но телу питательные вещества, лишь у голотурий она выполняет и функцию газообмена. На конце стенки кишки имеются кровеносные лакуны или, как у голотурий и морских ежей, хорошо развитые спинной и брюшной кровеносные сосуды, способные периодически сокращаться, что вызывает слабое движение «крови». Пульсации, обычной для кровеносной системы, у иглокожих нет, но имеется пульсация перикардия, стенки которого, как и у хордовых животных, образуют впячивание — сердце. Пульсация перикардия вызывает слабое движение жидкости, однако правильного движения «крови» у иглокожих пет, да и газового обмена, столь характерного для кровеносных систем, у них, за исключением голотурии, не установлено. Функцию газового обмена выполняет жидкость полости тела.
Очень тесное отношение к нервной и кровеносной системе имеет система каналов, выстланных эпителием и развивающихся из обособленных участков общей вторичной полости тела. Эта система каналов называется псевдогемальной или перигемальиой. Она состоит из кольца, идущего вокруг глотки или пищевода, расположенного между амбулакральным кольцом и околоротовым нервным кольцом, и пяти радиальных каналов, проникающих в амбулакры и проходящих между радиальными нервами и амбулакральными каналами. Жидкость псевдогемальной системы одинакова по составу с жидкостью вторичной полости тела, содержит большое число целомоцптпв, а близкое расположение этой системы к нервной дает возможность предположить, что она служит не только опорой для кровеносной системы, как думали раньше, но и для питания нервных тяжей и предохранения их от сдавливания. Псевдогемальная система развита неодинаково у разных классов иглокожих. Так. у морских ежен и голотурий имеются лишь радиальные каналы этой системы, а у морских лилии она вообще, как правило, отсутствует.
Радиальный план строении иглокожих проявляется в части и в расположении половых органов. Половая система состоит из полового тяжа и половых желез, образующихся в качестве выростов первого. Наиболее своеобразное и примитивное строение имеет половая система морских лилий. У них половые тяжи в виде пяти плотных лент проходят в лучи и ветвятся, повторяя разветвления последних. Концевые веточки их заходят в особые выросты на лучах, называемые пиннула-ми, и принимают вид полых слепых мешков. Половые продукты развиваются из клеток половых мешков внутри пиннул и выводятся наружу через разрывы их стенок. У морских звезд, офиур и морских ежей половой тяж имеет вид пятиугольника, стенки которого, разрастаясь, дают начало половым железам, расположенным интеррадиаль-по. Половые железы у морских звезд имеют вид пяти пар ветвящихся гроздевидных мешочков, помещающихся в основании лучей или заходящих в них далеко. Они открываются наружу короткими каналами между лучами. У офиур многочисленные мелкие половые железы расположены на внутренней стороне пяти пар особых выводковых сумок (бурс) и открываются непосредственно в эти сумки, которые сообщаются с внешней средой через узкие щели, находящиеся на ротовой стороне диска в основании лучей. У морских ежей пять гроздевидных половых желез помещаются на верхней части панциря, причем у многих неправильных морских ежей пятимерное строение нарушается исчезновением задней железы. У голотурий же имеется единственная половая железа, состоящая из пучка простых или ветвящихся трубочек. Все трубочки соединяются в один половой проток, который у большинства форм открывается наружу в передней части тела, в спинном интеррадиусе.
Нарушение пятплучевой симметрии у современных иглокожих встречается довольно часто. Так, все голотурии и неправильные морские ежи являются билатеральносимметричными животными с хорошо выраженными передним и задним концами тела. Да и у всех остальных иглокожих, кроме некоторых многолучевых морских звезд, ветвистых офиур и лилий, у которых наблюдается увеличение числа каменистых каналов и мадрепоро-вых пластинок, фактически можно провести только одну плоскость симметрии, песмотря на радиальное строение, из-за положения осевого комплекса органов, расположенного лишь в одном интеррадиусе. Эта черта современных иглокожих унаследована ими от билатеральносимметричных предков, тогда как двусторонняя симметрия голотурий и неправильных морских ежей скорее должна рассматриваться как вторичное явление, связанное с утратой радиальной симметрии в связи с переходом их к ползающему и роющему образу жизни.
О с е в о й к о м н л е к с о р г а н о в очень характерен для строения иглокожих и представляет собой пространственное объединение весьма разнообразных органов. Он образован амбулакральной, кровеносной и половой системами, а также участками вторичной полости тела. Основными составными частями этого комплекса являются каменистый канал, мадрепоровая пластинка с поровыми каналами, осевой орган, идущий вдоль каменистого канала в виде продолговатого мешка, два обособленных участка вторичной полости тела (левый и правый осевые синусы), заключающие в себе осевой орган, и, наконец, половой тяж, помещенный в особом участке полости тела — половом синусе. Осевой орган образован сплетением кровеносных сосудов, отходящих от ротового кровеносного кольца, перикардием (правый осевой синус) и значительным количеством соединительной ткани, а также подвижными клетками — амебоцитами. Функции осевого комплекса довольно многообразны. С помощью пульсирующего перикардия вызывается движение жидкости в кровеносной системе; путем притока и оттока морской воды через мадрепоровую пластинку происходит регулирование гидростатического давления в амбулакральной системе и удаление ненужных веществ. Осевой орган служит, с одной стороны, местом накапливания и выведения из организма продуктов распада, с другой — лимфатической железой, образующей амебоциты — клетки, свободно передвигающиеся в жидкости полости тела. Возможно, что осевой орган, связанный с половым тяжом, является также органом внутренней секреции. Осевой комплекс органов имеется у морских звезд, морских ежей и офиур, хотя его положение и соотношение органов в пем не одинаковы у разных классов в связи со смещением мадрепорита. У морских лилий и голотурий осевого комплекса органов нет, а имеются лишь каменистые и норовые каналы, которые очень часто открываются непосредственно в общую полость тела.
Целом (вторичная полость тела) у большинства иглокожих достигает значительного объема, выстлан ресничным эпителием, заключает в себе кишечник и большую часть внутренностей, покрытых этим эпителием. Лишь у морских лилий в связи с развитием значительного количества сое-динительпотканных мезодермальных тяжей, заполняющих полость тела, да у ряда офиур, у которых имеются особые эктодермальные сумки (бурсы), глубоко вдающиеся в полость тела, объем его резко уменьшается. Вторичная полость тела возникает из двух задних целомов личинки, причем мезентерий. разделяющий правый и левый целомы личинки, у многих взрослых иглокожих сильно редуцируется и сохраняется только у голотурий, следуя за изгибами кишечника. Полость тела заполнена полостной, или перивисцералыюй, жидкостью, очень прозрачной и близкой по составу к морской воде, по содержащей примесь белков н большое количество клеточных элементов. Эта полостная жидкость находится в постоянном движении, вызванном ресничным эпителием, выстилающим полость тела, и играет очень важную роль в процессах обмена веществ. Клеточные элементы (амебоциты), содержащиеся в пери висцеральной жидкости, участвуют в распределении питательных веществ и выделении продуктов распада.
Полостная жидкость имеет существенное значение и в процессах дыхания: у некоторых иглокожих в перивисцеральной жидкости имеются клетки с гемоглобином или другими дыхательными пигментами; кроме того, многие органы дыхания образованы стенками полости тела. Так, наружные околоротовые, сильно разветвленные (перистомальные) жабры большинства морских ежей представляют собой выпячивание обособленного участка вторичной полости тела — ротового, или перифарингеального, целома, расположенного вокруг пищевода. Через стенки жабр, нокрытых чрезвычайно тонким слоем кожи, растворенный в воде кислород проникает в полостную жидкость. Кожные жабры, или папулы, в виде тонкостенных сократимых очень мелких пузырьков имеются у морских звезд. Наряду с жабрами существенную роль в дыхании, очевидно, играют амбулакральные ножки и другие тонкостенные придатки амбулакральной системы, так как у ряда форм в клетках жидкости амбулакральной системы найден пигмент, близкий к гемоглобину. Большинство иглокожих весьма нуждается в кислороде и быстро погибает в плохо вентилируемых аквариумах. Однако органы дыхания у них развиты слабо и химизм дыхания изучен еще мало. Настоящие органы дыхания — водные легкие — имеются только у голотурий. У тех голотурий, у которых водные легкие отсутствуют, дыхательную фунцию, очевидно, могут выполнять ротовые щупальца, кишечник, сравнительно тонкая кожа тела и, наконец, спинные придатки амбулакральной системы, через которые проникает кислород. У офиур дыхание происходит через тонкостенные мешковидные камеры — бурсы, расположенные в основании лучей и периодически заполняющиеся морской водой благодаря сокращениям спинной стенки диска и наполнениям и опорожнениям желудка. Физиология дыхания иглокожих изучена недостаточно, но вполне вероятно, что различные системы органов могут принимать участие в дыхательных процессах наряду с выполнением ими других функций.
Органы пищеварения развиты неодинаково в разных классах иглокожих, что связано с различием в способах питания и с различным планом строения представителей отдельных классов.
Пищеварительпая система начинается ртом, который в зависимости от способов питания может быть снабжен скелетом или лишен его. У голотурий рот, окруженный щупальцами, помогающими при ловле добычи, переходит, как у морских ежей и лилий, сначала в короткую эктодермальную глотку, а затем в длинный узкий эптодермальный кишечник, длина которого значительно превышает длину тела животного. Кишечник поэтому образует петли, лежащие во вторичной полости тела и подвешенные к стенке тела при помощи мезентериальных тяжей. У голотурий, морских ежей и морских лилий можно различить передний, средний и задний отделы кишечника. Задний конец кишки открывается наружу выводным (анальным) отверстием, которое у морских лилий расположено вблизи рта, у морских ежей — на стороне, противоположной ротовой, или на краю скорлупы. У голотурий задняя кишка расширяется в довольно объемистую мускулистую клоаку, которая у большинства видов открывается наружу на заднем конце тела. У офиур и ряда морских звезд глотка отсутствует и рот ведет непосредственно в объемистый мешковидный желудок. прикрепленный к стенке тела мезентериаль-ными тяжами. Желудок офиур складчатой формы, с радиальными п интеррадиальными выростами, заканчивается слепо, так как офиуры не имеют ни печеночных придатков, ни задней кишки, ни анального отверстия. У морских звезд желудок имеет 5 пар печеночных выпячиваний, заходящих в лучи и обильно выделяющих пищеварительные соки. Таким образом, кишечный каиал иглокожих сохраняет особенности, унаследованные современными животными от двустороннесимметрич-ных предков. Он почти целиком развивается из первичного кишечника и не имеет радиального строения. Влияние пятилучевой симметрии сказывается только па кишечнике морских звезд, обладающем пятью парами печеночных отростков.
Процессы пищеварения иглокожих изучены недостаточно. Считают, что пищеварение у них внеклеточное. кроме внутриклеточного переваривания жиров. В стенках кишечника и его эпителии имеется много амебоцитов, участвующих в пищеварении. В кишечном соке найдены ферменты: липаза, амилаза, инверт^за, трипаза, бутираза, протениаза и некоторые другие. В эпителии кишечника находятся железистые клетки (слизистые. зернистые, секреторные), содержащие полисахариды. гликоген, липоиды.
Мало изучены и процессы выделения иглокожими продуктов распада, особенно процессы выделения жидких веществ. Специальных органов выделения, кроме урночек у некоторых голотурий, у иглокожих нет, а функцию выделения выполняют многочисленные целомоцнты полости тела, кровеносной системы, осевого органа, амбулакральной системы и ресничные перитонеальные клетки, поэтому выделение носит диффузный характер. В одном случае целомоцнты, нагруженные твердыми продуктами распада, выбрасываются наружу через разрывы сгенок тела, причем избираются места с наиболее юнкпмн покровами (папулы, перистомальные ????ры. амбулакральные ножки, водные легкие ???? п т. д.). В другом случае целомоцнты ??? с продуктами распада откладываются в соеденительной ткани в разных местах тела, образуя крупные окрашенные массы. Химическая природа твердых и жидких продуктов распада изучена недостаточно, однако у иглокожих установлено наличие кератина и кератинина, которые характерны, кроме них, только для позвоночных животных.
Развитие иглокожих всегда сопровождается сложным превращением с прохождением стадии пелагической двубокой личинки. У иглокожих всех классов развитие на ранних стадиях протекает более или менее сходно, но затем наблюдаются значительные различия. У большинства форм размножение только половое, яйца откладываются в воду, где они оплодотворяются и развиваются. После дробления, которое, как правило, полное, радиального типа, образуется типичная бластула, покрытая ресничками. На одном полюсе бластулы происходит глубокое впячивание (инвагинация), дающее начало энтодермальной средней кишке, и бластула превращается в гаструлу. Первое важное изменение в строении гаструлм происходит с образованием по бокам первичной кишки выпячиваний целомических карманов. Порядок их образования весьма изменчив у различных личинок, но наиболее обычным и первичным является независимое образование трех пар левых и правых целомических мешочков путем деления одного непарного мешка, отшнурованного от первичной кишки. Три левых целома (передний, средний и задний) значительно крупнее трех правых целомов (редуцированных), что нарушает двустороннюю симметрию личинки. В дальнейшем, в отличие от личинок червей, членистоногих и моллюсков, первичный рот, или бластонор, у личинок иглокожих превращается в анальное отверстие, а ротовое отверстие образуется вторично путем впячивания эктодермы брюшной стороны навстречу изгибающемуся концу первичной кишки. Подобное образование рта свойственно также погонофорам, полухордовым и хордовым животным, что сближает их с иглокожими и позволяет отнести последних к группе вторичноротых животных.
После смещения анального отверстия на брюшную сторону личинка иглокожих становится вполне двусторониесимметричной, с выпуклой спинной и вдавленной брюшной стороной, она обладает околоротовым венчиком ресничек и тремя парами целомических мешков. Эта стадия развития личинок называется д и п л с в р у л о й и является общей для большинства иглокожих. При дальнейшем развитии днплеврула испытывает значительные изменения, разные в разных классах иглокожих, проявляющиеся особенно в видоизменении внешней формы и формы околоротового венчика ресничек. Меньше всего изменяется форма диплев-рулы у голотурий. Их личиночная стадия называется а у р и к у л я р и е й. Она имеет овальную форму, рот. лежащий в глубине брюшного вдавления и окруженный одним венчиком мерцательных ресничек. а также скелет в виде мелких микроскопических телец. Личинки морских звезд называются б и п и н и ария м н, офиур — о ф и о и л у т е у с а м и, а морских ежей — э х и и о п л у т е у с а м и. Все названные личинки характеризуются присутствием на геле особых шпуров, снабженных ресничками, при помощи которых они могут свободно плавать. Личника морских лилий — д о л и о л я р и я обличается от остальных личинок тем, что лишена рта и имеет бочопкообразное тело, опоясанное ресничным шпуром не сплошным, а расположенным в виде пяти ресничных колец. В дальнейшем все лилии проходят еще стадию личинки типа и е н т а к р и п ус. прикрепленной стеблем к субстрату.
Размеры личинок иглокожих очень незначительы, длина их менее 1 мм. Но среди них имеются и гиганты. Так, бипнпнарпи морской звезды ;m\ui луидии досшгает длины 1л мм. Период жизни личинок в планктоне весьма различен и определяется не только видом животного, по и окружающими условиями. Так, ??орме личинки одного и того же вида при размножении летом находятся в планктоне от 5 до 7 недель, а в более холодное время зтот период удлиняется. Срок пребывания личинок в планктоне зависит и от типа личинки. Личинки, развившиеся из яиц, бедных желтком (планктонотрофные личинки), проводя в планктоне от нескольких педель до 2—о месяцев, активно питаясь одноклеточными водорослями. Личинки, развившиеся из яиц, богатых желтком (лецитотрофные личинки), имеют ускоренное развитие и находятся в планктоне or нескольких дней до месяца. В процессе превращения во взрослое животное, лечинки иглокижнх претерпевают значительные изменениия. У морских ежей, морских звезд и офиур большая часть тела личинки и ее органов разрушается и погибает; у голотурий атрофируется лишь небольшая передняя часть личинки, а остальная часть переходит в тело взрослой особи. Растут иглокожие очень медленно, в течение всей жизни, половой зрелости достигают через несколько лет. Рассмотренные выше типы личинок являются основными для различных классов иглокожих, однако следует заметить, что внешний вид личинок и способ их превращения во взрослый организм неодинаковы у разных видов животных этого типа.
Находясь в планктоне, личинки могут перемещаться на большие расстояния от места обитания взрослых форм. Это, с одной стороны, положительный факт, способствующий расселению вида, с другой стороны, как планктонные организмы, личинки иглокожих поедаются другими животными. Выедание компенсируется продуцированием иглокожими чрезвычайно большого количества яиц, чем обеспечивается выживание вида.
Очень часто иглокожие проявляют заботу о потомстве. В таких случаях мать прикрывает телом отложенные яйца или вынашивает их. Обычно молодь вынашивается на определенных местах материнского организма, реже ползает по всему телу. У иглокожих известны случаи живорождения, когда зародыш развивается внутри материнского организма, помещаясь в полости тела либо в половых трубках, либо в бурсальных выпячиваниях (у офиур) и получает от него питание, необходимое для развития. Живорождение особенно распространено у арктических и антарктических видов. В этом случае развитие прямое, без стадии планктонной личинки.
Наряду с половым размножением у иглокожих в отдельных случаях наблюдается и бесполое размножение, когда тело взрослой особи делится пополам или на несколько частей. Каждая образовавшаяся часть восстанавливает недостающие участки. Такой способ размножения наблюдается у некоторых морских звезд, офиур и отдельных голотурий.
В известной связи с бесполым размножением стоит весьма характерное для иглокожих явление регенерации, т. е. способность восстановления утраченных отдельных органов пли частей тела. Очень часто при неблагоприятных условиях или при нападении врага иглокожие отбрасывают (автотомируют) лучи, выбрасывают внутренности, а иногда даже распадаются на части. Скорость восстановления утраченных частей различна и зависит от температуры. Так, у тропических форм и в летнее время регенерация протекает быстрее, чем у арктических п бореальиых форм. Восстановление утраченных частей у молодых особей идет быстрее, чем у старых, однако личинки иглокожих почти не сиособны к регенерации. Это полезное защитное приспособление в очень слабой степени развито у морских ежой. Тело их покрыто прочным панцирем, предохраняющим от внешних повреждений, поэтому морские ежи могут при неблагоприятных условиях отбрасывать и затем восстанавливать только педицеллярии, иглы и амбулакральные ножки.
Относительно продолжительности жизни иглокожих имеется мало сведений, однако вполне вероятно, что живут они долго. На пластинках скелета некоторых иглокожих установлены годичные кольца роста. По подсчетам этих колец возраст крупных морских ежей исчисляется 35 годами, однако некоторые зоологи считают, что эта цифра завышена и средний возраст морских ежей — 10 или 15 лет. Примерно столько же живут офиуры. Некоторые морские звезды только к 14-летнему возрасту достигают своих обычных размеров. Максимальный возраст морских лилий около 30 лет. Голотурии, возможно, живут 5 лет или немного больше.
Все иглокожие обитают исключительно в морях и весьма чувствительны к малейшему опреснению воды. Они отсутствуют в Каспийском море, в Балтийском представлены всего 3 видами, а в Черном — только 8 видами, зато в Баренцевом, Карском, Чукотском и Охотском морях составляют основную часть донных животных. Сравнительно немногие виды способны переносить довольно значительные понижения солености — до 15—24°/00 — и даже иногда встречаются в эстуарных участках; другие виды прекрасно чувствуют себя в воде с чрезвычайно высокой соленостью. Примером может служить Красное море, обладающее соленостью 43—47 г солей на 1 кг воды и обильно населенное самыми разнообразными иглокожими. Иглокожие живут в морях и океанах всех широт земного шара, на самых разнообразных грунтах и глубинах — от приливо-отливной зоны до максимальных глубин океанических желобов, которые бывают глубже 10 ООО м. Чаще иглокожие приурочены к определенным глубинам и встречаются либо исключительно па литорали, либо в сублиторальной или батиальной зоне, либо обитают только в абиссальной зоне, причем в пределах отдельных групп иглокожих видовое разнообразие иногда увеличивается с глубиной. Однако среди иглокожих значительное число видов оказываются эврибатпыми1 формами, диапазон вертикального распространения которых иногда превышает 5000—7000 м. Таких видов, которые могут встречаться на самых различных глубинах, особенно много среди офиур. Для ряда иглокожих были отмечены сезонные миграции из глубин к берегам и, в связи с размножением или изменениями температуры, от берега на большие глубины.
Степень приуроченности иглокожпх к определенным грунтам различна. Многие голотурии, морские ежи и офиуры живут исключительно па илистом грунте или песке, но некоторые представители этих классов предпочитают каменистый грунт. Однако вполне вероятно, что характер грунта не играет существенной роли в распространении иглокожих. Более важную роль в их расселении играют соленость, глубина, температура, наличие течений, история развития морей и океанов, а также целый ряд биологических факторов. Среди иглокожих встречаются виды, предпочитающие определенную температуру. Так, арктические формы встречаются только при температу-рых, близких к О СС, а высокоарктические — исключительно при отрицательных температурах, тогда как арктическо-бореальные виды являются более эвритермными, т. е. могут переносить значительные колебания температуры. Среди бореаль-ных видов также имеются эвритермные формы, которые даже зимой остаются на литорали, в условиях снежного покрова, но основная масса видов с наступлением холодов уходит в более глубокие воды. Особенно пышно представлены иглокожие в фауне тропических морей и прибрежных зон, что связано с более постоянными высокими температурами и с наиболее разнообразными условиями обитания в этих районах по сравнению с холодными или умеренными морями или с большими глубинами. Тем не менее даже и в последних двух районах иглокожие довольно многочисленны и нередко принимают участие в образовании определенных сообществ, или биоценозов, являясь руководящими формами большинства из них. Богата иглокожими фауна Тихого океана, тогда как фауна Атлантического — заметно беднее. Некоторые виды иглокожих имеют довольно широкое распространение по всем океанам, другие обладают разорванными ареалами (биполярные, амфибореальные и амфипацифические виды), третьи приурочены только к тропическим, бореальным, арктическим или антарктическим областям. Многие виды, роды и семейства иглокожих эндемичны для определенных районов, они прекрасно характеризуют как прибрежные, так и глубоководные фауны различных областей. Поэтому зоогеографическое значение иглокожих весьма велико.
Иглокожие играют важную роль и в общей экономике моря, являясь многочисленными и типичными его обитателяхми. Подобно моллюскам и другим животным, они участвуют в поддержании определенного солевого состава морской воды, потребляя в процессе питания большое количество солей, особенно карбоната кальция, из которого строят свой скелет. Иглокожие обладают способностью извлекать пз воды радиоактивные вещества, которые накапливаются прежде всего у них в жидкости полости тела. Кроме того, иглокожие служат пищей различным животным. Например, треска, пикша, камбала, зубатка и другие рыбы поедают значительное количество офиур и морских ежей. Морских ежей употребляет в пищу и морская выдра — калан. Охотятся за иглокожими и некоторые крупные моллюски и ракообразные. С другой стороны, мпогие иглокожие являются хищниками и сами поедают различных животных. Особенно хищными слывут морские звезды, которые могут съедать рыбу, превышающую размерами их тело, и часто нападают на устричные банки и рыболовные сети. Некоторые иглокожие используются в пищу человеком. Среди них наиболее важное значение имеют съедобные голотурии — трепанги, которых насчитывается около 40 видов и разновидностей. Наиболее старыми потребителями трепангов являются жители Китая, Японии, Малайского архипелага. Однако и в этих странах трепанги скорее лакомое блюдо, так как высокая цена и ограниченная продуктивность мест лова препятствуют более широкому потреблению этих животных. В Китае трепангов по целебным свойствам сравнивают с корнем женьшеня и называют «корнем моря». Кроме голотурий, в Японии и по берегам Средиземного моря добывают отдельные виды морских ежей, икру которых используют в пищу. Иглокожие являются прекрасным объектом для экспериментальных эмбриологических исследований, и в последние годы их яйца, особенно яйца морских ежей, использовались в космических полетах для выявления влияния излучения на живые организмы. Полезными для человека являются и ископаемые иглокожие, так как некоторые строительные материалы, такие, как дербширский, бельгийский мрамор, трахитовый известняк, состоят главным образом из остатков данных животных. Кроме того, ряд ископаемых иглокожих, особенно морских ежей и морских лилий, используют в качестве руководящих форм, важных для стратиграфии.
Почти все иглокожие ведут свободный образ жизни, паразитов среди них, можно сказать, нет, хотя питание молоди некоторых видов офиур па колониях кораллов и последующее переселение их на диск и в полость тела взрослых особей собственного вида, а также обитание карликовых самцов нескольких видов офиур постоянно около ротового отверстия самки и высасывание ими с помощью специально заложенных в глотке мышц пищевых соков изо рта самки, пожалуй, можно подвести под понятие паразитизма. Комменсалы среди иглокожих известны давно. Часто офиуры поселяются между иглами морских ежей, где. очевидно, находят себе пищу и защиту, а представители одного из родов тропических офиур постоянно живут на ротовой поверхности морских лилий около амбулакральных борозд и пользуются нищей, собираемой этими животными, не причиняя хозяевам заметного вреда. Целый ряд других животных — простейших, гидроидов, ракообразных, моллюсков и даже рыб — являются комменсалами иглокожих и обитают на них, прикрепившись к иглам, поверхности тела морских ежей или морских звезд, либо поселяются в полости тела, кишечнике и других органах различных иглокожих, делясь с ними пищей и получая от них некоторую защиту. Однако существуют и настоящие паразиты иглокожих. Правда, они немногочисленны и относятся к разным группам животных. Очень распространенными и наиболее древними паразитами иглокожих, известными еще с каменноугольного периода, являются кольчатые черви подкласса Myzostomida. Объектом интенсивного заражения этими паразитами становятся морские лилии, как более или мепее прикрепленные животные. Часто в голотуриях паразитируют некоторые моллюски, а в офиурах — различные паразитические ракообразные и неполовозрелая форма трематоды, являющейся во взрослом состоянии паразитом зубатки.
Среди современных иглокожих лишь стебельчатые морские лилии неподвижно прикреплены к субстрату, а все « стальные животные свободно передвигаются. (Обычно иглокожие ползают но грунту как но горизонтальным, так и по вертикальным поверхностям, реже зарываются в грунт, и лишь единичные виды голотурий ведут пелагический образ жизни н могут встречаться в толще воды. Закапывание в грунт распространено довольно широко во всех группах иглокожих, кроме морских лилий, у которых в связи со своеобразным способом питания амбулакральные борозды должны быть всегда чистыми. Большинство форм зарывается г. груиг па небольшую глубину, оставляя часть тела пли часть лучей над грунтом. Закапывание в грунт у многих иглокожих связано с питанием. Они заглатывают грунт без разбора, причем захват грунта с находящимися в нем питательными веществами происходит механически вместе с поступательными движениями животных. Грунтоеды встречаются среди многих голотурий, морских ежен и некоторых морских звезд. Большинство иглокожих питается животной пищей, но некоторые из них потребляют исключительно прибрежные водоросли или даже остатки наземной растительности.
По поводу происхождения иглокожих и филогенетических взаимоотношений внутри этого типа до енх пор существуют только многочисленные догадки и гипотезы. Уто объясняется до некоторой степени тем, что появление иглокожих относится к глубокой древности, так как в конце кембрия и в начале ордовика, свыше 500 млн. лет назад, уже имелись представители почти всех известных нам классов иглокожих. Предполагают, что иглокожие возникли в докембрии и наиболее древние из них. вероятно, имели слаборазвитый скелет, что не позволило им сохраниться в ископаемом состоянии.
На основании данных эмбриологии, сравнительной анатомии и палеонтологии считают, что предки иглокожих, очевидно, имели двусторонне-спммстрнчное. вытянутое тело с пятью парами щупалец, трубковидным кишечником и тремя нарами целомов. Рот у них находился иа передней части тела, анус — па задней. Эти животные вели прпдоппый образ жизни, который вызвал некоторую асимметрию в их строении. Наиболее близкими к такому предку были представители примитивных классов гомалозой и пельматозой. Однако гомалозой не имели признаков радиальной симметрии и были очень обособлены от остальных иглокожих. У примитивных пельматозой (ранне-кембрийских эокриноидей и ордовикских паракри-нопдей) уже был намечен переход от двусторонней симметрии к радиальной. Прикрепление к субстрату правой стороной переднего конца тела животного, которое имело место у представителей этого подтипа, вызвало смещение рта влево и вверх, ануса — вправо и приобретение радиального плана строения. Таким образом, эволюция дельматозой шла в направлении развития радиальной симметрии, и у высших представителей этого подтипа, ряда морских лилий, у которых наблюдается увеличение числа каменистых каналов, она охватывает почти всю их.организацию, за исключением кишечника и способа отхождения приводящих бороздок от рта. Среди современного подкласса Arliculata стебельчатые морские лилии представляют собой реликты древней фауны, а наиболее развитыми и процветающими в настоящее время являются бесстебельчатые морские лилии отряда Comatulida.
Относительно происхождения подтипа эхинозой единого мнения нет. Включенные в него классы столь различны, что выведение их из общего эхпнозойного ствола вместе с геликоплакоидеями и объединение морских ежей с голотуриями вызывает много возражений. Прежде всего, веретено-видные геликонлакоидеи так своеобразны, что связать их с каким-либо классом этого подтипа просто невозможно. Голотурии же, кроме некоторых черт анатомического строения, ни по данным эмбриологии, пи по биохимическим данным, ни но общему типу строения не могут быть сближены с морскими ежами. Личинки голотурий весьма отличны от личинок морских ежей, но обнаруживают сходство с личинками морских звезд, да и в биохимии мышечного сокращения у голотурий и у морских звезд играет роль аргипин-фосфаген, а у морских ежей, как и у офиур — креатин-фос-фагеи. Па основании этого скорее можно думать о наличии общего прототипа у этих четырех классов, которые ранее относились к одному подтипу элеутерозой.
Наличие у голотурий ряда примитивных черт, таких, как- непарная половая железа, положение полового отверстия и мадрепорита вблизи рта, развитие с переходом личинки голотурии без особых изменений в тело взрослого животного с сохранением личиночного скелета и становление нятплучевой симметрии амбулакральной системы у некоторых форм через трехлучевую, свидетельствует о древности этого подкласса. Благодаря слабому развитию скелета палеонтологическая история юлотурпй мало известна. Наиболее примитивным считают отряд голотурий сдревовидными щупальцами (Dendrochirola).
Что касается класса морских ежей, то такие признаки, как наличие осевого комплекса органов и полового тяжа, некоторые особенности развития скелета ротовой стороны сближают их с морскими звездами и офиурами. О сходстве с офиурамн свидетельствуют морфология личинок и некоторые биохимические свойства. Однако имеющийся лишь у морских ежей аристотелев фонарь, характерные только для них амбулакральные пластинки. пронизанные порами для прохождения амбулакральных ножек, тело без выступающих лучей, образованное главным образом за счет акти-нальной стороны, не дают возможности предположить о близком родстве морских ежей с морскими звездами и офиурами. Ответвление морских ежей от общего ствола произошло, вероятно, тогда, когда классы морских звезд и офиур еще формировались. Раньше предков морских ежей, морских звезд и офиур видели в формах, близких к ранним эдриоастероидеям, но это не подтвердилось. Основной группой ствола морских ежей, от которой могли произойти все остальные ежи, считают отряд Lepidocentroida. От него, вероятно, берут начало копьеносные ежи (Cidaroida). Современные кожистые ежи (Echinothuriidae) столь сходны с палеозойскими лепидоцентроидами, что большинство зоологов помещают их в этот отряд. Напротив, Дарем и М е л ь в и л ь, которые выделяют всех палеозойских морских ежей и современных цидароид в подкласс Perischoechinoidca, а всех мезозойских и остальных современных ежей в подкласс Euechinoidea, рассматривают эхинотуриид в качестве самостоятельного отряда подкласса эуэхиноидей и считают, что они вместе с остальными постпалеозойскими морскими ежами произошли от цидароидных предков. Д. М. Ф е-дотов полагает, что наиболее древние правильные постпалеозойскне морские ежи отряда Аи1о-donla, представители которого имели тонкий, нередко гибкий панцирь, гораздо легче выводятся из морфологически разнообразной группы лепи-доцентроид, чем из более специализированной группы цидароид. Предполагают, что с отрядом Aulodonta связан происхождением отряд Sli-rodonta, а отряд Canianxlonla произошел от предков из отряда Stirodonla. Эволюция правильных морских ежей, кроме группы цидароид. шла по пути создания прочного панциря, развития ам-булакров и амбулакральных ножек, образования нз простых амбулакральных пластинок сложных, со многими норами и т. д. Считают, что неправильные морские ежи в виде двух ветвей отдели лнсь от одной из групп постпалеозойскпх правильных морских ежей. Морские ежи отряда 11о-lectypoida по ряду признаков близки к формам отрядов Stirodonla или Aulodonla и могли быть родоначальной группой для других неправильных морских ежей. Эволюция неправильных морских ежей шла в сторону развития двусторонней симметрии в связи с переходом пх к роющему образу жизни.
Предположение Ф е л л а о близких родственных связях между морскими лилиями и формами подтипа Asterozoa необоснованно, так как некоторое сходство в строении скелета морских лилий и сомастероидей чисто поверхностное. Оно отчасти наблюдается лини» у современных бесстебельчатых морских лилий и ордовикски\ сомастероидей и отсутствует у палеозойских морских лилий и сомастеропдсй. Сомастероидеи могли быть предками морских звезд и офиур, но связать их с ка-кой-либой группой пельматозой невозможно. Древние морские звезды и офиуры обладали рядом общих черт и имели общих предков. В настоящее время это четко отграниченные классы. Эволюция морских звезд шла в сторону облегчения скелета, увеличения размеров тела, удлинения лучей, перехода от пассивного питания к хищничеству. Уже в палеозое появились формы, которые были близки к современным.
Офиуры прошли более сложный путь развития. В палеозое существовали офиуроподобиые формы, сильно отличающиеся от современных. Эволюция офиур была связана с переходом к движеппю с помощью рук (подвижных лучей) вместо движения при помощи амбулакральных ножек, как у морских звезд. Это привело к обособлению рук от диска, замыканию амбулакральных борозд, превращению парных амбулакральных пластинок в позвонки рук, развитию специальной мускулатуры рук и системы наружных покровных пластинок. Типичные офиуры известны с мезозоя, и в настоящее время это наиболее многочисленный по числу видов класс среди всех иглокожих.
Тип иглокожих включает 3 подтипа: стебельчатые иглокожие (Pelmatozoa), охинозои (Echinozoa) и астерозои (Asterozoa).
Иглокожие (лат. Echinodermata) — тип исключительно морских донных животных, большей частью свободноживущих, реже сидячих, встречающихся на любых глубинах Мирового океана. Насчитывается около 7000 современных видов (в России — 400). Наряду с хордовыми, иглокожие относятся к ветви вторичноротых животных (Deuterostomia). Современными представителями типа являются морские звёзды, морские ежи, офиуры (змеехвостки), голотурии (морские огурцы) и морские лилии. В состав этого типа входят также приблизительно 13 000 вымерших видов, которые процветали в морях, начиная с раннего кембрия. / (Википедия)
Книги:
 Жизнь животных. Том 2. Моллюски. Иглокожие. Членистоногие / В. Е. Соколов. — 1988
Жизнь животных. Том 2. Моллюски. Иглокожие. Членистоногие / В. Е. Соколов. — 1988
 Зоология беспозвоночных в двух томах. Том 2: от артропод до иглокожих и хордовых / Вестхайде В., Ригер Р — 2008 с
Зоология беспозвоночных в двух томах. Том 2: от артропод до иглокожих и хордовых / Вестхайде В., Ригер Р — 2008 с
 Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989
Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989