

ОПИСАНИЕ:
Класс Euglenophyceae Schoenichen in Eyfurth & Schoenichen, 1925
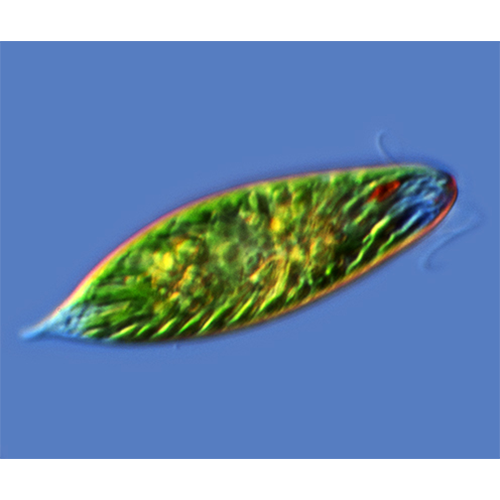
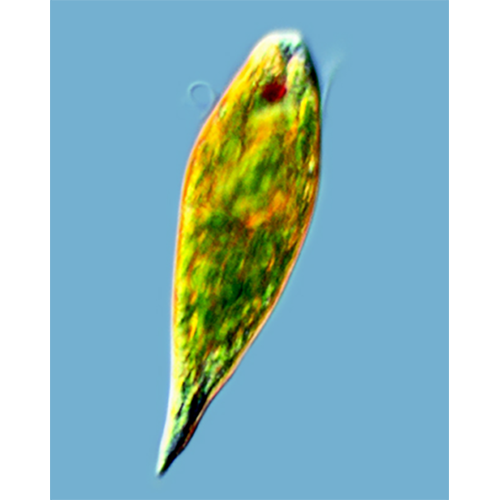
В составе класса одиночные и колониальные, фототрофные (зеленые) и гетеротрофные (бесцветные) протисты. Форма тела широко варьирует от овальной, веретеновидной до листовидной и игловидной. Длина тела 5—500 мкм и более. У большинства видов 2 гетероконтных гетеродинамичных жгутика, выходящих из дна жгутикового резервуара на переднем конце тела. Жгутики с параксиальным тяжем, простыми мастигонемами, длинной переходной зоной и 3 микротрубочковыми корешками. Поверхность тела покрыта кутикулой, некоторые виды имеют домик. Движение жгутиковое и метаболическое (эвгленоидное).Фототрофные виды имеют фоторецептор, включающий стигму и парафлагеллярное тельце. Хлоропласты в числе от 1 до нескольких сотен, с оболочкой из 3 мембран. Тилакоиды в ламелле собраны по 3. Основные пигменты хлоропластов: хлорофилл а, b и fl- каротин. Имеются пиреноиды. Митохондрии с пластинчатыми кристами. Способы питания: голофитное, сапрозойное, голозойное, миксотрофное. Гетеротрофные и некоторые фотосинтезирующие виды имеют цитостом. Основные резервные вещества — парамилон и липиды.
Эвглениды одноядерны. Хромосомы конденсированы на протяжении всего жизненного цикла, при делении ядра ядрышко не разрушается. Размножение агамное путем продольного бинарного деления. В жизненном цикле обычны пальмеллевидные формы и цисты. Класс объединяет около 1000 свободноживущих, паразитических и эпибионтньгх видов, которые распространены всесветно.
ИСТОРИЯ ИЗУЧЕНИЯ
Первое описание ряда основных видов и родов эвгленовых принадлежит Эренбергу (Ehrenberg, 1830). К их числу относятся: представители родов Euglena, Astasia, Distigma, Colacium, Trachelomonas. Фауна их интенсивно изучалась и в последующие годы (Dujardin, 1841; Perty, 1852; Stein, 1878; Klebs, 1883, и др.), что позволило выделить эту группу в отдельный отряд в типе Protozoa. Накопление знаний по морфологии и физиологии способствовало обоснованию растительной природы эвгленовых.Идея о выделении их в качестве одной из групп одноклеточных жгутиковых водорослей принадлежит Ценковскому (1870). Эвгленовые были включены в систему споровых растений в качестве отдельного типа (отдела) Euglenophyta Butschli, 1884 с 1, обоснованным позже, порядком (классом) Euglenales Popova, 1951. В настоящее время доминирует идея о первичности гетеротрофных форм в эволюции эвгленовых. Подавляющее большинство бесцветных эвгленид исходно не имеют хлоропластов, которые, по-видимому, были приобретены позднее в результате симбиоза.
В современной системе протистов эвгленовые вместе с кинетопластидами выделены в особый тип Euglenozoa Cavalier-Smith, 1981 и представляют в нем отдельный класс Euglenoidea Butschli, 1884 (Cavalier-Smith, 1978; Corliss, 1984).
В настоящее время эвгленовые известны в качестве одной из важнейших групп протистов, имеющих большое значение в природе, а также как объект для целого ряда экспериментальных работ. Поэтому в течение последних десятилетий много внимания уделено изучению их фауны, описанию новых видов, родов, семейств, издан ряд хорошо иллюстрированных пособий, атласов, определителей (Св1ренко, 1938; Goidics, 1953; Попова, 1955, 1966;Mignot, 1966; Leedale, 1967, 1978, 1985; Асаул-Ветрова, 1975; Попова, Сафонова, 1976; Bourrelly, 1985; Willey et al., 1988; Triemer, Ott, 1990; Dawson, Walne, 1994; Лихачёв, 1997; Simpson, 1997). Бьщо сделано подробное описание группы эндопаразитических эвгленовых из кишечника, полости тела и яйцевых мешков пресноводных веслоногих рачков (Michajlow, 1972, 1978).
Большие успехи достигнуты в культивировании ряда видов эвгленовых, получены их аксенические (безбактериальные) культуры (Pringsheim, 1946; Гол- лербах, Полянский В. И., 1951; Wolken, 1961, и др.). Массовые клональные культуры (особенно эвглен и астазий) широко применяются в самых разнообразных исследованиях — морфологических, электронно-микроскопических, биохимических, генетических, молекулярно-биологических, а также в биотехнологии (Epstein, Allaway, 1967, 1968; Leedale, 1967; Buetow, 1968, 1982; Зайцева, Шанина, 1976; Суханова, 1983, 1985). В течение всех последних лет подробно изучается значение эвгленовых в природе и их экология (Винберг, 1966; Sladecek, 1973; Anderson, 1987; Larsen, Patterson, 1990; Simpson et al., 1996/97).
МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
Внешнее строение
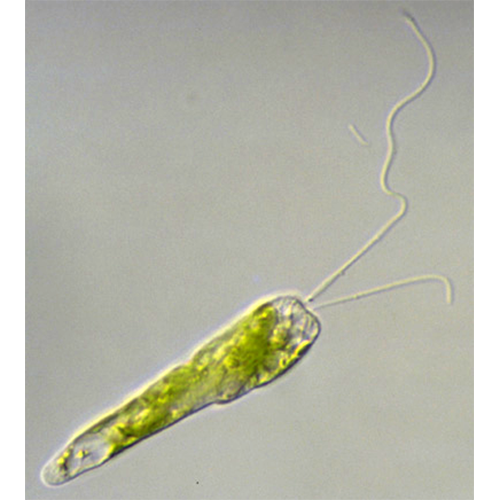
Форма и размеры тела. Форма тела трофозоитов широко варьирует: она может быть шаровидной, эллипсоидальной, листовидной, яйцевидной, игловидной, веретеновидной. Последняя наиболее типична и широко распространена в разных таксонах класса. Передний конец тела более или менее закруглен, задний же часто вытянут и заканчивается заостренным отростком. Одну из особенностей эвгленовых составляет спиральная скрученность тела, выраженная неодинаково у разных видов, влияющая на форму тела и обусловливающая спиральную симметрию у ряда видов.Жгутиковый аппарат. Большинство видов имеет 2 неравных по длине, гетеродинамичных жгутика, и только у немногих, например у эвтрепций, оба Ж1уги- ка практически одинаковы. Есть также виды с 1 или несколькими жгугиками.
Жгутики выходят наружу через отверстие, расположенное на переднем конце тела. Оно ведет в узкий канал, который продолжается в углубление грушевидной формы — жгутиковый резервуар. Выходящие наружу жгутики несут длинные мастигонемы, расположенные в 1 ряд с одной стороны, а также короткие мастигонемы, расположенные спирально по всей длине ундулиподии. Обязательная внутренняя структура жгутиков — параксиальный тяж. В длинной переходной зоне жгутиков часто отсутствует даже поперечная пластинка и только у Entosiphon обнаружена мощная псевдозвездчатая структура.
2 кинетосомы расположены параллельно друг другу; от них отходят корешки, состоящие из микротрубочек.
Все 3 корешка направлены в переднюю часть клетки и проходят вдоль стенок жгугикового резервуара (Farmer, Triemer, 1988), поддерживая его форму. У некоторых видов в системе корешков имеются фибриллярные элементы.
Жгутиковый резервуар эвгленовых отличается крупными размерами и достаточно сложным упорядоченным расположением связанных с ним органелл. Он выстлан плазмалеммой, которая представляет собой продолжение плазмалеммы поверхности тела. Жгутики отходят от дна резервуара, и длинный двигательный жгутик проходит вдоль дорсальной стенки резервуара. Второй, у большинства видов короткий жгутик, часто не выходящий за пределы резервуара, расположен вблизи вентральной его стенки.
Сократительная вакуоль расположена в брюшной стенке жгутикового резервуара, в который изливается ее содержимое при сокращении. У фотосинтезирующих эвглен (род Euglena) недавно обнаружен рудиментарный цитостом, который, вероятно, частично сохраняет функциональную активность (как пиноци- тозный канал), так как в этом месте обнаруживаются лизосомы. Он открывается в дистальную часть резервуара. Скрытый цитостом эвглен гомологичен цитостому бесцветных видов эвгленовых и кинетопластид (Willey, Wibel, 1985; Surek, Melkonian, 1986).
Среди эвгленовых есть ряд хищных гетеротрофных видов, у которых развиты весьма сходные приспособления для захвата пищевых организмов. Это представители родов Peranema, Heteronema, Entosiphon, обладающие глоточным палочковым аппаратом. У Р. trichophorum глоточный аппарат лежит в вентральной части переднего конца клетки. Его стенки укреплены 2 трихитами — глоточными палочками. Трихиты состоят из многочисленных микротрубочек, ассоциированных с фибриллярными структурами. Весь этот ротоглоточный комплекс обеспечивает заглатывание как мелких, так и крупных пищевых объектов.
Глоточный палочковый аппарат сходного строения имеют Heteronema acus и другие виды этого рода, а также виды рода Entosiphon. У Е. sulcatus глоточные структуры проходят почти по всей длине тела. Они образованы тремя мощными пучками микротрубочек, с проходящими вдоль них фибриллярными «парусами», обращенными во внутреннее пространство глотки (Triemer, Fritz, 1986). Пищей этих хищных эвгленовых жгутиконосцев служат не только мелкие, но и крупные простейшие, их цисты, а также мелкие многоклеточные.
С жгутиковым аппаратом фототрофных эвгленид тесно связан фоторецептор, состоящий из парафлагел- лярного тельца и стигмы. Парафлагеллярное тельце представляет собой расширенный участок длинного (дорсального) жгугика и расположено у его основания в жгутиковом резервуаре. Напротив параф- лагеллярного тельца, вблизи дорсальной стенки жгутикового резервуара, расположена стигма (глазок). Эта органелла состоит из отдельных глобул, каждая из которых покрыта мембраной. Стигма окрашена либо в красный, либо в оранжево-красный цвет. Это связано с тем, что содержимое каждой глобулы представлено каплей липида, в которой растворен бета-каротин. Механизм взаимодействия стигмы и парафлагел- лярного тельца окончательно не выяснен, но есть наблюдения, свидетельствующие о том, что если стигма затеняет парафлагеллярное тельце, то эвглена изменяет направление своего движения. В последнее время в прилегающем к стигме участке плазмалеммы обнаружен родопсин, что, вероятно, поможет в ближайшем будущем выяснить ее функцию.
Движение. Эвгленовым свойственны 2 типа движения: жгутиковое и метаболическое (эвгленоидное).
С помощью жгутика они передвигаются в водной среде, вращаясь вокруг своей продольной оси. Метаболическое движение, или метаболия, выражается в удлинении и сужении, укорочении и расширении тела или отдельных его частей, свертывании, способности изгибаться в стороны. Для метаболии характерны также перистальтические волны, хорошо выраженные у Eutreptia, Distigma, у некоторых видов рода Parastasia. Волны сокращений тела проходят вдоль его продольной оси. Каждая волна начинается с расширения заднего конца и перемещается к переднему, вызывая продвижение тела вперед. Затем жгутиконосец восстанавливает свою прежнюю форму тела, после чего происходит перемещение цитоплазмы спереди назад.
Метаболия свойственна видам, обитающим на поверхности субстрата и ведущим ползающий образ жизни. У таких видов жгутик может отсутствовать совсем или недоразвит. Переход к метаболическому движению происходит и в тех случаях, когда эвглены отбрасывают жгутик при каких-либо воздействиях. У астазий, эвтрепций, дистигм жгутиковое и метаболическое движения осуществляются одновременно (Попова, 1966). Поверхность тела эвгленовых покрыта слизью, которая значительно облегчает метаболическое движение.
Среди эвгленовых есть и неметаболичные виды, обладающие плотной оболочкой, которая препятствует сокращениям тела. Это Euglena acus, виды родов Cyclidiopsis, Rhabdomonas и другие. Степень выраженности метаболии (слабая, сильная), а также ее отсутствие используются в качестве одного из таксономических признаков.
Поверхностные структуры. Поверхностные структуры эвгленовых представлены кутикулой, часто с сильно развитым слоем гликокаликса. Под плазма- леммой находятся эластичные лентовидные пластинки белковой природы, расположенные спирально вокруг тела(Mignot, 1966). Эти пластинки рельефно выступают под плазмалеммой, образуя на поверхности клетки продольные или спирально закрученные «штрихи», или ребра.
Между пластинками у большинства видов эвгленовых есть промежутки, которые обусловливают эластичность, гибкость поверхности тела, необходимую для осуществления метаболических сокращений. В промежутках между белковыми пластинками, а также под ними располагаются микротрубочки, которые связаны между собой мостиками, обеспечивающими скольжение микротрубочек относительно друг друга. По-видимому, этот механизм обусловливает эвгленоидное движение.
Поверхность тела всех видов Trachelomonas и Strombomonas защищена особым образованием — домиком. Форма домиков широко варьирует от яйцевидной, шаровидной, цилиндрической до веретеновидной. Домик может быть гладким или покрытым шипами, бугорками, различными выростами, порами, сетевидными утолщениями, На переднем конце домик имеет горлышко с отверстием для выхода двигательного жгутика, а на заднем конце у некоторых видов имеется хвостовидный отросток.
Форма и размеры домиков, их орнаментация специфичны для вида и являются важными таксономическими признаками. Стенки домиков представляют собой утолщенный поверхностный слой, состоящий из уплотненных слизистых веществ полисахаридной и белковой природы. Целлюлозы в стенках домиков нет, они инкрустированы солями железа, которые придают им характерный бурый или коричневый цвет разных оттенков, делают их более жесткими и прочными.
Цитоплазма
Хлоропласты. Фотосинтезирующие виды эвгленовых имеют хлоропласты, которых нет у бесцветных гетеротрофных видов. Однако у некоторых из них, например у Khawkinea quartana, обнаружены рудиментарные пластиды. Число хлоропластов у разных видов варьирует от 1—2 до нескольких сотен. Если хлоропластов немного, то число их постоянно для вида и может служить таксономическим признаком.Форма хлоропластов разнообразна. Они могут быть дисковидными, звездчатыми или лентовидными. Оболочка хлоропластов состоит из 3 мембран. Крупные хлоропласты несут пиреноиды, у мелких они отмечены не всегда. На поверхности пиреноидов, обращенной к цитоплазме, откладывается парамилон, синтез которого происходит вне хлоропластов.
Хлоропласты травянисто-зеленого цвета, содержат хлорофиллы а и b, β-каротин, диадиноксантин и другие пигменты в малом количестве. Тилакоиды в ламелле группируются по 3.
Митохондрии. Эти многочисленные органеллы расположены в цитоплазме всех участков клетки. Им свойственны пластинчатые кристы. Эвглениды, обитающие в анаэробных условиях, лишены митохондрий. Это продемонстрировано недавно на примере Postgaardi mariagerensis (Simpson et al., 1996/1997).
Сократительная вакуоль. Чаще всего сократительная вакуоль 1, и расположена она в передней части тела возле дорсальной стенки жгутикового резервуара. Содержимое вакуоли изливается в полость жгутикового резервуара, а из него — наружу.
Аппарат Гольджи. Диктиосомы имеют типичное строение, и число их неодинаково у разных видов. Часто они представлены стопками из 10—15 цистерн.
Резервные вещества. Основное резервное вещество, специфичное для всех видов эвгленид, — полисахарид парамилон. Центрами его синтеза в клетке служат пиреноиды, однако он образуется и у бесцветных форм. По своей химической природе парамилон представляет 0 - 1,3-связанный глюкан, который откладывается в цитоплазме в виде зерен различной величины и формы, специфичных для отдельных родов и видов. У одних видов зерна парамилона мелкие и заполняют всю цитоплазму, например у Euglena gracilis, Astasia longa, Parastasia fennica и других; у ряда видов кроме мелких зерен формируются еще 1—4 крупных зерна, называемых парамилиями. Место расположения парамилий в цитоплазме строго постоянно и специфично для вида. Форма парамилей может быть палочковидной, как например у Cyclidiopsis acus, Euglena acus или кольцевидной, что характерно для родов Phacus, Lepocinclis. Парамилии свойственны слабометаболичным видам и неметаболичным видам с постоянной формой тела.
У бесцветных эвгленовых кроме парамилона синтезируется гликоген и небольшие его запасы откладываются в цитоплазме в виде мелких зерен. Резервным веществом служит, также нейтральный жир, который откладывается в цитоплазме в виде капель. У некоторых эвглен (Euglena rubra) в качестве запасного вещества отмечен гематохром, который накапливается в цитоплазме при температуре 27—30 °C и дает клетке красную окраску.
Ядро
Эвгленовые — одноядерные организмы. Ядро чаще всего шаровидное, расположено либо в центральной части клетки, либо несколько смещено в нижнюю ее половину. В центре ядра находится крупное ядрышко. Хромосомы концентрируются в периферической зоне ядра. В интерфазном ядре хромосомы имеют высокую степень конденсации, и по своей ультраструктуре они сходны с митотическими хромосомами (Райков, 1978). Деление ядра осуществляется по типу закрытого плевро- или ортомитоза.Размножение
Основной формой размножения служит бинарное деление, но известна и палинтомия, свойственная всем известным видам Parastasia и некоторым представителям Khawkinea. Перед делением ядро обычно мигрирует к переднему концу клетки. Цитокинез у всех эвгленид происходит в продольном направлении, и борозда, разделяющая 2 дочерние особи, проходит от переднего конца тела к заднему. Фототрофным видам свойствен циркадный ритм делений. Бесцветные виды делятся асинхронно в течение всего времени суток. Деление может происходить в подвижном состоянии или в состоянии пальмеллы. При этом делящаяся особь выделяет большое количество слизи, которая образует прозрачную и довольно толстую оболочку.Жизненные циклы
В жизненных циклах свободноживущих и прикрепленных эвгленовых имеется несколько фаз: активные трофозоиты, делящиеся особи, пальмеллевидные формы и цисты покоя. В природных условиях в течение теплых сезонов эти фазы периодически сменяют друг друга, но зимой большинство эвгленовых находятся в состоянии цист покоя.У метаболичных видов цисты покоя имеют шаровидную форму и покрыты плотной оболочкой мукополисахаридной природы. У видов, не способных к метаболическим движениям (представители родов Cyclidiopsis, Phacus и др.), переход в покоящееся. состояние осуществляется просто путем отбрасывания жгутика. Изменений формы тела при этом не происходит.
Из немногочисленной группы эндопаразитических эвгленовых жизненные циклы достаточно подробно изучены у нескольких видов рода Parastasia (Michajlow, 1972, 1978; Вита, Суханова, 1983; Лихачёв, 1989). Все известные виды парастазий имеют сложные и довольно однотипные жизненные циклы, сходные с жизненным циклом Parastasia fennica. Он включает в себя 2 фазы: трофическую, проходящую в организме хозяина (рачка), и репродуктивную, протекающую в воде. На репродуктивной стадии образуются жгутиковые инвазионные клетки, похожие на свободноживущих астазий. Регулярное чередование трофической и репродуктивной фаз жизненного цикла происходит только в течение весны, лета и ранней осени; зимуют парастазии в организме хозяина или (в редких случаях) в цистах покоя.
ЭКОЛОГИЯ И ПРАКТИЧЕСКОЕ ЗНАЧЕНИЕ
Распространение и места обитания
Эвгленовые распространены всесветно. Огромное большинство их — обитатели пресных континентальных водоемов, и, вероятно, нет такого водоема, в котором нельзя было бы найти те или иные виды этих протистов. В шельфовых зонах морей и океанов преобладают гетеротрофные эвглениды (Patterson, Simpson, 1996).Популяции эвгленовых наиболее многочисленны в мелководных стоячих пресных водоемах лесной и лесостепной зон. Это водоемы, в которых много растительности, хорошо прогревается вода и содержится много органических веществ. Многочисленны эвгленовые и в удобряемых рыбоводных прудах, на рисовых полях, а также в биологических прудах, где происходит очистка сточных вод. В таких водоемах они часто представляют доминирующую группу организмов микропланктона и микробентоса. Развиваясь в массовом количестве, могут служить причиной зеленого и красного «цветения» воды.
Среди фотосинтезирующих эвгленид преобладают одиночные свободноживущие виды, которые ведут плавающий образ жизни: эвглены, факусы, трахеломонасы и многие другие. Напротив, гетеротрофные эвглениды большую часть времени ведут ползающий образ жизни, лишь ненадолго переходя к плаванию в поисках скоплений микроорганизмов, которые служат им пищей. Таковы, в частности, Petalomonas, Anisonema, Peranema, Ploeotia и Другие.
Небольшой группе эвгленовых свойствен неподвижный (на большей части жизненного цикла), прикрепленный образ жизни. Сюда относится несколько видов рода Colacium — эпибионтов ракообразных и других планктонных животных. К внешним покровам животных они прикрепляются передним концом тела с помощью слизистого стебелька или подушечки, образуя ветвистые колонии. Cyclidiopsis acus способны временно прикрепляться к какому-либо субстрату с помощью слизи, выделяемой ими на заднем, заостренном конце тела. В прикрепленном состоянии они вегетируют, размножаются и могут образовывать временные колонии.
Еще одну группу эвгленовых представляют эндопаразитические виды родов Astasia и Parastasia, обитающие в период трофической фазы жизненного цикла во внутренних органах пресноводных Copepo- da (Michajlow, 1972; Лихачёв, 1989).
Влияние факторов внешней среды
Пищевой фактор. Несмотря на то что эвглено- вым присущи все способы питания, свойственные фототрофным и гетеротрофным организмам, пищевой фактор, и прежде всего содержание в воде органических веществ, лимитирует их распределение по биотопам и распространение. Способность многих видов к миксотрофному или сапрофитному питанию обусловливает массовое развитие их в водоемах или в тех зонах водоемов, которые богаты органическими веществами. Поэтому большинство эвгленид может быть использовано в качестве индикаторных организмов водоемов с повышенной степенью сап- робности (Sladecek, 1973). Пищевыми объектами фаготрофных эвгленовых (роды Peranema, Urceolus и др.) служат бактерии, мелкие простейшие, водоросли.Температура. Популяции эвгленовых начинают развиваться ранней весной, и их численность увеличивается с повышением температуры в июне. В середине лета численность популяций снижается, но в конце лета и начале осени вновь значительно увеличивается. При этом время весеннего и осеннего максимумов развития различается в разных климатических зонах (Попова, 1966, и др.).
Снижение температуры воды осенью стимулирует формирование цист покоя, которые переживают в грунте водоемов зиму. Однако ряд холодолюбивых видов остается в активном состоянии поздней осенью и зимой подо льдом при температуре воды 4—6 °C. Один из холодолюбивых видов — Euglena viridis — образует многочисленные популяции поздней осенью в сильно эвтрофированных мелких водоемах, а также в биологических прудах. Популяции этого вида эвглен достигают численности до 1—3 и более миллионов особей в 1 л воды. Они локализуются в поверхностном слое, где активно осуществляют фотосинтез, улучшая кислородный режим биологических прудов и природных водоемов, что особенно важно для процессов минерализации органических веществ (Вин- берг, 1966; Попова, 1966; Суханова, 1983).
Холодолюбивые Euglena viridis, Е. proxima, трахеломонасы, перанемы и другие зимой могут вмерзать в лед, не теряя при этом своей жизнеспособности.
Интересные данные о полиморфизме вида по температурному оптимуму размножения получены для 26 штаммов Е. gracilis из разных мест обитания (Pringsheim, 1958). В пределах этих штаммов выделены 3 группы, различающиеся по температурному оптимуму размножения.
1. Эвглены прекращают размножение при 27 °C, остаются зелеными, и у них сохраняется стигма.
2. Эвглены интенсивно размножаются при 34 °C, но многие из них становятся бесцветными, теряя при этом стигму.
3. Эвглены интенсивно размножаются при 34 °C, остаются зелеными, и у них сохраняется стигма.
Данные этих экспериментов показывают, что в природе существуют разные физиологические группировки (расы) в пределах вида Е. gracilis, адаптированные к разным температурным условиям.
Свет. Продолжительность фаз клеточного и жизненного циклов, время размножения и другие жизненные функции фототрофных эвгленовых лимитируются наряду с температурой световым днем. Размножение этих организмов происходит в темноте.
При отсутствии света многие зеленые виды становятся бесцветными, гетеротрофными и нуждаются в органическом источнике углерода. Их морфология и клеточный метаболизм претерпевают глубокие изменения. У обесцвеченных в темноте эвглен (Ё. gracilis) уменьшается содержание каротиноидов, резорбируются пиреноиды, в цитоплазме откладываются большие запасы парамилона, у многих особей исчезает стигма. Обесцвеченные эвглены морфологически сходны с Astasia longa, что послужило причиной многих дискуссий о систематическом положении этого вида (Pringsheim, 1958; Попова, 1966). Перемещение обесцвеченных эвглен из темноты на свет сопряжено с восстановлением хлоропластов и других органелл, а также с восстановлением фотосинтетической активности. Однако некоторые особи после возвращения их из темноты в условия нормального освещения остаются бесцветными в течение длительного времени. Культивирование. Многие виды эвгленид легко культивируются в лабораторных условиях и живут в культуре в течение продолжительных сроков. Для культур разработано большое количество разнообразных органических и минеральных сред (Pringsheim, 1955; Wolken, 1961; Попова, 1966; Сухарева-Немакова, Каленик, 1977; Сухарева, 1989, и мн. др.). Большинство культуральных сред содержит минеральную основу, к которой добавляются различные органические вещества и витамины. Существуют и твердые среды, содержащие агар-агар, на которых живут многие виды в состоянии пальмеллы.
Е. gracilis образует в культурах настолько массовые популяции, что может быть использована в качестве корма для мальков рыб и других животных, а также как тест-объект для выяснения действия целого ряда лекарственных препаратов, например антибиотиков и витаминов.
Особенно большое научное и практическое значение имеют аксенические культуры Е. gracilis и Astasia longa, которые способны жить в культуральных условиях в течение длительного времени. Они служат объектами для многочисленных морфологических, физиологических, биохимических, молекулярно-биологических и генетических исследований, а в последнее время и для целей биотехнологии (Сухарева, 1989, и др.). Происхождение и эволюция. Вопрос о происхождении эвгленид изучается до сих пор. Сведений об их ископаемых формах очень мало, и они не вполне достоверны (Попова, 1966). Эвглениды представляют четко очерченную по своим морфологическим особенностям группу, в которой фотосинтезирующие и гетеротрофные виды обнаруживают высокую степень сходства по строению клетки. Однако нельзя отрицать сходства эвгленид, особенно фотосинтезирующих видов, и с одноклеточными водорослями (Зайцева, Шанина, 1976; Buetow, 1982; Рейвн и др., 1990).
Новые данные о наличии цитостома у фотосинтезирующих эвгленид и сравнительный анализ строения корешковых систем жгутиков с таковым кинетопластид позволили выявить гомологию этих структур в обеих группах. Кроме того, эвглениды и кинетопластиды имеют и другие черты сходства: ундулиподии жгутиков снабжены у тех и других параксиальными тяжами, образованными гомологичными белками, деление ядра осуществляется посредством закрытого митоза. Все эти признаки могут служить доказательством филогенетических связей между обеими группами и подтверждают происхождение фотосинтезирующих видов эвгленовых от бесцветных гетеротрофных форм (Taylor, 1976; Leedale, 1978; Willey, Wibel, 1985; Surek, Melkonian, 1986).
Книги:
 Протисты. Руководство по зоологии. Часть 1. Алимов А.Ф. 2000
Протисты. Руководство по зоологии. Часть 1. Алимов А.Ф. 2000
 Протисты. Руководство по зоологии. Часть 2. Алимов А.Ф. 2007
Протисты. Руководство по зоологии. Часть 2. Алимов А.Ф. 2007
 Протисты. Руководство по зоологии. Часть 3. Алимов А.Ф. 2011
Протисты. Руководство по зоологии. Часть 3. Алимов А.Ф. 2011
 Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987
Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987



 Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
 Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423
Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423
 Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989
Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989