
ОПИСАНИЕ:
Тип Cryptophyta Pascher, 1914 // Cryptista Cavalier-Smith, 1989 - Криптофитовые водоросли, Криптомонады
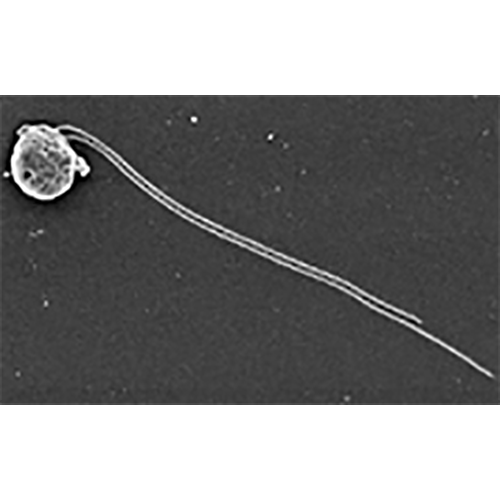
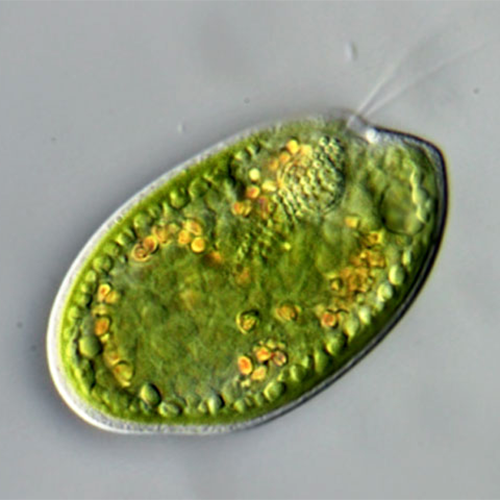
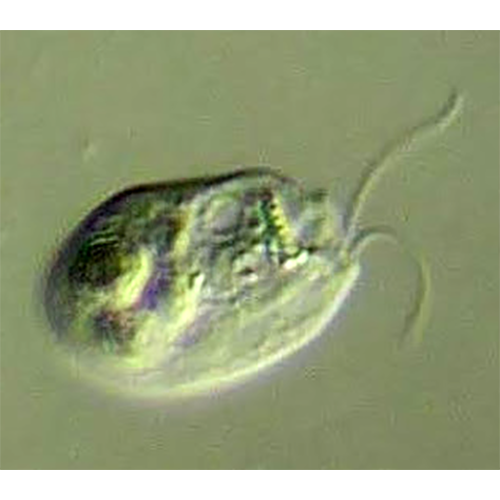
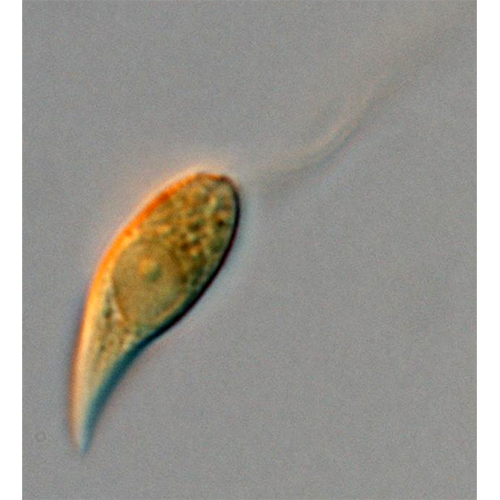
Одноклеточные подвижные организмы с типичным дорсовент- ральным строением. Имеют овальную, яйцевидную, почковидную или бобовидную форму. Тело клетки сжато с боков и имеет выпуклую дорсальную и плоскую (или вогнутую) вентральную сторону, передний край обычно косо срезан. Здесь расположена вестибулярная зона, от которой вдоль тела клетки отходит косая борозда; у начала борозды расположено отверстие глотки, уходящей внутрь тела на различную глубину. В вестибулярной зоне находятся основания 2 жгутиков, из которых один несколько длиннее другого. Глотка выстлана рядами крупных стрекательных органелл — эжектосом. Преимущественно строгие фототрофы, но есть и гетеротрофные формы, способные заглатывать и переваривать клетки бактерий. Клетки фототрофных крйптомонад могут иметь различную окраску — коричневую, сине-зеленую или красную за счет различного содержания фотосинтетических пигментов, таких как хлорофиллы а и с2, водорастворимые фикобилиновые пигменты фикоцианин и фикоэритрин. Хлоропласты криптомонад имеют оболочку из 4 мембран, из которых наружная является продолжением ядерной мембраны. Между 2 наружными и 2 внутренними мембранами находится перипластидное пространство, в котором располагаются рудиментарное ядро — нуклеоморф, цитоплазма с рибосомами и запасные питательные вещества. Половой процесс неизвестен. Одни формы всегда подвижны, даже при делении, другие образуют скопления заключенных в слизь неподвижных клеток. В редких случаях формируются круглые неподвижные цисты.История изучения
Первым ученым, исследовавшим криптомонад, был Эренберг (Ehrenberg, 1830, 1831), описавший 6 фототрофных видов, отнесенных к предложенному им роду Cryptomoiias, и 1 гетеротрофный вид, Chilo- manas paramaecium. Описания Эренберга, как обычно в те времена, были очень краткими и носили самый общий характер. В дальнейшем новые роды и виды описали Пашер (Pascher, 1913, 1914), Скуйя (Skuja, 1948, 1956) и другие. Подробный обзор морских форм сделал Батчер (Butcher, 1967). Разными авторами было описано более 200 видов криптомонад. Однако детали строения сложно организованных клеток крйптомонад плохо различимы в световом микроскопе, а описания многих таксонов, приводимые в ботанической литературе, являются недостаточно четкими. Многие признаки весьма изменчивы, без изучения культур установить природу организма затруднительно. Поэтому неудивительно, что в течение последнего десятилетия в результате использования электронного микроскопа систематика криптомонад претерпела резкие изменения. Большое значение для развития современной систематики криптомонад имели работы Санторе (Santore, 1984, 1985, 1987), хотя не всегда его взгляды получали признание. Переописания ряда родов и описания новых таксонов крйптомонад осуществили Хилл и Ветерби (Hill, 1991; Hill, Wetherbee, 1988, 1989, 1990). Эти описания основаны преимущественно на данных электронной микроскопии, но они иногда недостаточно конкретны и неоднозначны, поэтому и в наши дни определение родовой, не говоря уже о видовой, принадлежности этих организмов представляет значительные трудности. Поставив под сомнение правомерность использования ультраструктурных критериев в таксономии, Новарино и Лукас (Novarino, Lukas, 1993) пересмотрели систему крйптомонад, значительно сократив число родов. Создание коллекций типовых культур позволило в последнее время провести сравнение последовательностей генов 18S рРНК и обосновать выдвинутую ранее гипотезу о происхождении криптофит в результате симбиоза гетеротрофных двухжгутиковых монад и коккоидных красных водорослей (Marin et al., 1998).Морфология, цитология, физиология
Как уже было отмечено, клетки криптомонад обладают дорсовентральным строением, дорсальная сторона обычно выпуклая, тогда как вентральная вогнутая или плоская. На вентральной стороне расположена вестибулярная зона.Вестибулярная зона представляет собой мелкое углубление поверхности. Здесь начинается борозда, идущая обычно вниз вдоль клетки, тут же расположено отверстие глотки. Наличие борозды и ее длина относительно тела клетки — признаки, имеющие значение для систематики на уровне рода. Борозда часто несколько загнута. У некоторых форм на нижнем конце борозда имеет небольшое округлое расширение — устьице, что также является признаком, используемым в современной систематике криптомонад. Иногда края борозды сомкнуты, но отверстие устьица может при этом сохраняться. На дорсальной стороне вестибулярной зоны обычно находится вестибулярный язычок. Язычки могут быть овальными, квадратными, плоскими, округлыми или заостренными. Язычок укреплен цитоскелетной пластинкой. Мешковидная глотка у разных форм имеет разную длину и проходит вдоль или поперек тела. Иногда она разветвляется. У некоторых форм глотка укреплена цитоскелетными образованиями, электронно-плотными исчерченными пластинами. Вопрос о функциях и значении глотки неясен. У большинства фототрофных форм никогда не удавалось наблюдать заглатывания каких-либо частиц.
Покровы клетки представлены перипластом, который состоит из внутреннего компонента, находящегося под плазмалеммой и представляющего собой либо сплошную пластину, либо совокупность многочисленных белковых пластинок различной формы (Brett et al., 1994). Внешний компонент перипласта находится снаружи на поверхности плазмалеммы и представлен пластинками, чешуйками или слизью. В зависимости от характера перипласта поверхность клетки может быть ровной или структурированной. Например, поверхность у Chroomonas состоит из прямоугольников, а у Rhinomonas — из шестигранников (Kugrens, Lee, 1987). Перипласт отсутствует в области глотки и вестибулума.
Во внутренний компонент перипласта обычно встроены многочисленные мелкие стрекательные органеллы, на срезах клеток криптомонад часто можно видеть пустые ямки, оставшиеся после выстреливания их содержимого. Глотка выстлана крупными стрекательными органеллами — эжектосомами, расположенными рядами; число рядов эжектосом иногда указывается в диагнозах родов и видов. Эжектосомы содержат скрученные рулоном ленты; при раздражении клетки ленты распрямляются и выстреливаются во внешнюю среду. Значение эжектосом остается неизвестным; нет и доказательств их защитной роли; однако удалось наблюдать стремительный ракетоподобный рывок клетки при выбросе из глотки освобожденных из эжектосом лент.
Интерфазное ядро, крупное, занимает обычно нижнюю часть клетки. Оно содержит ядрышко и скопления постоянно конденсированных хромосом.
Основания не равных по длине жгутиков находятся у дорсального края вестибулярной зоны возле отверстия глотки, иногда внутри глотки. Окончания жгутиков слегка заострены, на их поверхности располагаются различные придатки. У многих форм длинный жгутик снабжен 2 противоположными рядами трубчатых мастигонем (1—2 мкм длиной) с одиночной концевой нитью (0.5—1 мкм). У основания длинного жгутика находится вздутие, от которого отходит пучок длинных мастигонем. По-видимому, он образует защитную сетку у отверстия глотки. Короткий жгутик несет короткие мастигонемы (около 1 мкм длиной) с коротким и длинным волосками на концах. На поверхности мембраны жгутиков иногда обнаруживаются чешуйки — семиугольные розетки 140—170 нм в диаметре. Они могут покрывать также й тело криптомонады. В отличие от других групп протистов у криптомонад наблюдается сильное варьирование характера жгутиковых придатков. Приведенное выше описание справедливо лишь в отношении некоторых форм. Варьирует структура мастигонем, их расположение, кроме чешуек могут присутствовать иглоподобные крючковидные выросты или волоски. Систематическое значение подобных различий пока не установлено; во всяком случае, в современных диагнозах родов криптомонад эти признаки не упоминаются.
Особенностью переходной зоны жгутиков криптомонад является наличие здесь 2 септ, из которых верхняя находится на уровне плазмалеммы. Кинетосомы (базальные тела жгутиков) ассоциированы с фибриллярными и микротрубочковыми жгутиковыми корешками. У разных форм эти структуры различаются. Характерен для криптомонад ризостиль — лента микротрубочек, соединенных сократимыми фибриллами. Ризостиль может проходить в непосредственной близости от ядра и достигать заднего конца клетки. Возможно, что при этом он как бы стягивает клетку вдоль ее длинной оси, что обусловливает искривленную форму тела у видов Campylomonas. На поперечном срезе ризостиль образует изогнутую пластинку. Микротрубочки ризостиля иногда снабжены изогнутыми пластинками — крыльями и соединены с дополнительным фибриллярным тяжем. У многих форм отмечен исчерченный фибриллярный корешок, идущий вдоль поверхности митохондрии. Восходящий микротру- бочковый корешок соединяется с пластиной, образованной цитоскелетными микротрубочками.
Сократительная вакуоль находится у апикального конца клетки, она открывается в область глотки около ее начала. В передней части клетки расположен аппарат Гольджи, образованный 1—2 диктиосомами, состоящими из 12—15 цистерн.
Митохондрия, по-видимому, 1, сильно разветвленная, с пластинчатыми, ламеллярными кристами, образует периферическую и центральную сети; периферическая сеть расположена под плазмалеммой, центральная —соответственно в центре клетки.
Большинство криптомонад осуществляют фотосинтез и являются автотрофами. Редкие формы (например, Chroomonas pochmannii) миксотрофны, осуществляют фотосинтез и могут захватывать и переваривать клетки бактерий. Известно 2 рода гетеротрофных криптомонад — Goniomonas и Chilomo- nas. Бактерии обнаруживаются в пищеварительных вакуолях, но механизм их попадания в тело криптомонады до сих пор не описан.
Фототрофные формы содержат 1 или 2 хлоропласта. Иногда один из хлоропластов образует 2 большие лопасти, соединенные тонким мостиком, часто трудно установить, сколько именно этих органелл содержит клетка. В хлоропластах находятся специализированные плоские мембранные цистерны — тилакоиды, содержащие основные светособирающие пигменты и компоненты электрон-транспортных цепей. Тилакоиды обычно расположены попарно, иногда в группах по 3. Клетки фототрофных криптомонад могут иметь различную окраску — коричневую, сине-зеленую или красную за счет различного содержания фотосинтетических пигментов, таких как хлорофиллы а и с2, водорастворимые фикобилиновые пигменты фикоцианин и фикоэритрин.
Фикобилины состоят из а- и Р-субъединиц фико- билинпротеинов и линейных тетрапирролов — хромофоров. Эти пигменты расположены во внутритила- коидном пространстве и не образуют каких-либо видимых структур, подобных фикобилисомам красных и синезеленых водорослей. Каждый вид содержит только один из фикобилиновых пигментов, причем существует несколько типов фикоцианина и фикоэритрина, характеризующихся различными максимумами поглощения света (в описаниях родов максимум указан в нанометрах после названия пигмента). Интересно отметить, что генетические исследования свидетельствуют об отсутствии у криптомонад белков фикоцианинов. Предполагается, что замена хромофора у фикоэритрина мимикрирует спектроскопические характеристики истинных фикоцианинов (Marin et al., 1998). Представители одного рода обычно содержат один определенный пигмент, хотя вопрос о таксономическом значении пигментации у криптомонад является пока дискуссионным (Hill, Rowan, 1989). Нередко в хлоропласте находятся небольшие липидные глобулы.
В хлоропласте, в его выросте, или на стебельке над ним может располагаться пиреноид. Это белковое тело, содержащее основные ферменты фиксации углекислоты. Пиреноиды иногда рассечены мембранами тилакоидов. Крахмальная обкладка пиреноида не всегда хорошо выражена, что затрудняет его обнаружение в световом микроскопе. У немногих форм криптомонад в хлоропласте располагается глазок — стигма, которая состоит из группы липидных глобул на периферии хлоропласта, иногда в его выросте. Фототрофные формы способны к фототаксису. Их светочувствительность связывают с присутствием флавопротеинов на жгутиках.
Хлоропласт окружен 4 мембранами. 2 внешние мембраны принято обозначать как хлоропластный ЭПР. Внешняя мембрана хлоропластного ЭПР переходит во внешнююю мембрану ядра. Перипластидный компартмент, ограниченный хлоропластным ЭПР и 2 внутренними спаянными мембранами, включает зерна крахмала, нуклеоморф и цитоплазму с эукариотическими рибосомами.
Нуклеоморф — рудиментарное ядро эукариотического фототрофного симбионта — может быть ассоциирован с пиреноидом, ядром, иногда располагается в выросте хлоропласта, иногда — внутри пиреноида. Нуклеоморф обычно имеет округлую или грушевидную форму, окружен двойной мембраной, в которой обнаруживаются поры; он делится амитотически, простой перетяжкой. В нуклеоморфе выделены зоны скопления РНК, соответствующие зоне ядрышка, и зоны скопления ДНК. Геном нуклеоморфа у исследованных в этом отношении форм представлен 3 хромосомами и не является пассивным образованием; некоторые его гены работают, в частности гены 18S рРНК.
Клетка криптомонады состоит в отличие от большинства других протистов из 4 различных клеточных компартментов, содержащих собственный геном: цитоплазматического, перипластидного, пластидного и митохондриального, каждый из которых ограничен двойной мембраной.
Размножение, жизненный цикл
Размножение крйптомонад происходит путем продольного деления клетки, делящиеся клетки часто сохраняют подвижность. Митоз начинается с репликации кинетосом, ядро перемещается в передний конец клетки. Ядерная оболочка в профазе фрагментируется и частично разрушается, в случае открытого митоза из хлоропластного ретикулума формируется дополнительная перинуклеарная оболочка. ЦОМТами служат базальные тела, но на полюсах веретена располагаются исчерченные фибриллярные корешки и нуклеоморфы. Цитокинез идет параллельно кариокинезу, борозда деления формируется от переднего к заднему концу клетки. Каждая дочерняя клетка получает 1 материнскую кинетосому и 1, построенную de novo. Криптомонады — это гаплоидные организмы. Половой процесс неизвестен. Однако описан вид Proteomonas sulcata, имеющий популяции гаплоидных и диплоидных клеток, которые различаются по размерам, морфологий и организации жгутикового аппарата (Hill, Wetherbee, 1986). В редких случаях в жизненном цикле образуются круглые неподвижные цисты, при прорастании которых происходит деление их содержимого и из цисты выходят 2 вегетативные подвижные клетки.Экология
Криптомонады относятся к организмам-убиквистам. Они характеризуются значительной устойчивостью к загрязнению, встречаются даже в сточных водах и навозной жиже. Способны адаптироваться к резким изменениям условий среды: например, Chroo- monas africana не теряет жизнеспособности при переносе из морской воды в пресную. Многие виды являются морскими организмами, их разнообразие больше такового пресноводных. Последние могут быть найдены в планктоне. Особенно часто они населяют прибрежные участки водоемов, старицы, пруды и болота. Обычно число клеток в сообществах невелико, но иногда происходит массовое развитие криптомонад, они вызывают «цветение» воды, окрашивая ее в тот или иной цвет. Известны виды Cryptomonas, способные развиваться на поверхности снежного покрова и подо льдами Арктики. Хороший рост культур наблюдался при нейтральной или слабокислой реакции среды; сильный свет тормозит их рост.Криптофиты имеют большое значение в круговороте кислорода, углерода, азота и фосфора в водоёмах, в синтезе органики из минеральных веществ. Криптофиты служат пищей для различных представителей зоопланктона, которым, в свою очередь, питаются рыбы. Ветвистоусые рачки интенсивно питаются криптофитами в эвтрофных прудах в зимнее время. Однако цветение воды в прудах, обусловленное Cyanomonas, приводило к массовой гибели сомов.
В настоящее время криптофиты находят применение в исследовательской сфере. Так, недавно открытые родопсины водоросли Guillardia theta, представляющие собой анионные каналы, могут использоваться в качестве очень эффективного оптогенетического метода для подавления нейронов
Симбиотические связи
Криптофиты живут в качестве эндосимбионтов в клетках инфузории Myrionecta rubra. Эта инфузория может вызывать нетоксичное красное цветение воды в зоне апвеллинга. Цвет обусловлен красновато-коричневыми хлоропластами эндосимбионта. Криптофитовая водоросль, находящаяся внутри инфузории, окружена двойной мембраной (плазмалемма водоросли и пищеварительная вакуоль клетки-хозяина) и функционирует как полноценный автотроф. Ей присущи все особенности строения криптофитовых водорослей, но она лишена перипласта, жгутикового аппарата и эжектосом. Инфузория, содержащая эндосимбионтов-криптофит, целиком полагается на них в своём питании и утрачивает клеточный рот.Некоторые криптофиты являются эндосимбионтами некоторых динофлагеллят (например, Dinophysis[en]), причём в некоторых случаях у них может сохраняться ядро криптофитовой водоросли, а в других случаях сохраняются только пластиды
Филогения
В соответствии со схемой, предложенной Кюг- ренсом и Ли (Kugrens, Lee, 1991), исходными в эволюции морфологических типов крйптомонад считаются формы, близкие к представителям рода Goniomonas. У них вестибулярно-глоточное пространство не было разграничено и открывалось непосредственно в окружающую среду. К настоящему времени общепризнано, что пластиды криптомонад появились в результате вторичного эндосимбиоза, в котором эндобионт был фототрофным эукариотом. Этот этап эволюции связан с образованием устойчивого симбиоза с эукариотами (предположительно из группы Rhodophyta) и уменьшением размеров вестибулярного пространства. При этом пространство глотки, направленной внутрь клетки, сообщалось с окружающей средой через борозду. В дальнейшем происходили смыкание вентральных стенок вестибулярной зоны и редукция борозды. Глотка стала открываться в окружающую среду только через вестибулярную зону, как у представителей рода Chroomonas. Эжектосомы переместились с периферии вестибулярной зоны на мембраны, выстилающие глотку, и приобрели вертикальное расположение относительно длинной оси клетки. В процессе эволюции, по-видимому, произошли редукция хлоропласта и повторный переход к гетеротрофии, что привело к возникновению рода Chilomonas, представители которого обладают сильно редуцированной бороздой, развитой глоткой и лейкопластами. Приведенная гипотетическая схема подтвержается анализом нуклеотидных последовательностей генов 18S рРНК (Marin et al., 1998).Таким образом, криптомонады являются организмами-химерами. Только представители одного рода — Goniomonas, не обладающие ни хлоропластами, ни нуклеоморфом, возможно, являются потомками исходного организма-хозяина.
Книги:
 Протисты. Руководство по зоологии. Часть 1. Алимов А.Ф. 2000
Протисты. Руководство по зоологии. Часть 1. Алимов А.Ф. 2000
 Протисты. Руководство по зоологии. Часть 2. Алимов А.Ф. 2007
Протисты. Руководство по зоологии. Часть 2. Алимов А.Ф. 2007
 Протисты. Руководство по зоологии. Часть 3. Алимов А.Ф. 2011
Протисты. Руководство по зоологии. Часть 3. Алимов А.Ф. 2011
 Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987
Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987



 Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
 Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423
Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423
 Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989
Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989