
ОПИСАНИЕ:
Тип CILIOPHORA Doflein, 1901 — ИНФУЗОРИИ
Общая часть
Инфузории — ресничные простейшие, число свободноживущих видов которых в современной фауне достигает, по разным оценкам, от 3 до 30 тыс. (Finlay et al., 1996; Finlay, Fenchel, 1999; Foissner, 1999) и, как правило, принимается равным 8—9 тыс. видов (Согliss, 1979; Lynn, Corliss, 1991; Lynn, Small, 2000). Число эндокомменсалов и паразитов среди представителей типа Ciliophora обычно считается значительно меньшим (около 30 % от общего числа), хотя, как и общее количество, оно не определено более или менее точно (Finley et al., 1996; Foissner, 2001). Несмотря на значительный полиморфизм представителей типа, инфузории — одна из наиболее гомогенных групп в царстве Protista (Lynn, Corliss, 1991). Это преимущественно гетеротрофные формы, принадлежность которых к инфузориям определяется несколькими достаточно четкими признаками: наличием ядер двух типов — макро- и микронуклеусов (ядерный дуализм или гетероморфизм); присутствием на протяжении всего жизненного цикла или хотя бы на некоторых его стадиях поверхностно расположенных ресничек, образованных типичным набором микротрубочек (9+2) и связанных с базальными телами (кинетосомами); значительным развитием и специализацией поверхностного слоя клетки (кортекса), существенная составляющая которого — инфрацилиатура — система, образованная единичными, парными или множественными кинетосомами и их микротубулярными и микрофиламентоз- ными дериватами; прохождением полового процесса в виде конъюгации, в ходе которой партнеры, как правило, временно объединяются и обмениваются гаметиче- скими ядрами; присутствием у большинства представителей под поверхностью клетки специальных кортикальных пузырьков (альвеол), окруженных унитарной мембраной и подстилающих клеточную плазмалемму.Инфузории — преимущественно свободноплавающие, всесветно распространенные одиночные простейшие, хотя встречаются виды-эндемики и разнообразные прикрепленные и колониальные формы. Коммен- сальные (экто- и эндо-) и паразитические виды могут заселять различные компартменты тела многочисленных беспозвоночных и позвоночных животных, а также некоторых простейших.
Размер клетки инфузорий существенно варьирует, и среди представителей типа встречаются как действительно микроскопические клетки с линейными размерами, не превышающими 10 мкм (некоторые Colpodea и Oligohymenophorea), так и хорошо видимые простым глазом «гигантские» Spirostomum ambiguum и Bursaria truncatella, достигающие длины 3—4.5 мм. Соответственно клеточный объем инфузорий может колебаться от 102—103 мкм3 yNivaliella и Cyclidium до 108— 109 мкм3 у Bursaria и трофонтов Ichthyophthirius (Lynn, Corliss, 1991). Столь же сильно варьирует и форма тела инфузорий: если наиболее часто это простейшие сферические или вытянуто-овальные с разными модификациями, то среди обитателей псаммона, например, можйо встретить и дорсовентрально сплющенных червеобразных Karyorelictea, а в сапропеле — спирально скрученных представителей Armophorea и листообразных Loxodes.
Представители типа Ciliophora занимают самые разнообразные экологические ниши и, как уже было сказано, могут становиться экто- и эндокомменсалами и паразитами других одно- и многоклеточных животных. В свою очередь многие Ciliophora служат хозяевами для бактериальных экто- и эндобионтов, грибов, водорослей и других простейших. Свободноживущие формы встречаются повсеместно: в любых пресных, солоноватых и полносоленых морских водах как постоянного, так и временного характера. Пруды, реки, озера (в том числе соленые и гиперсоленые), моря, болота, лужи и заплески, песок, почва, лесная подстилка, листья и кора деревьев, тающий лед — вот далеко не полный перечень экологических ниш, занимаемых инфузориями. Очевидно, что распространение и число инфузорий, в целом, ограничивается (помимо присутствия некоторого количества воды) только температурой, которая должна быть выше точки замерзания, и присутствием пищи.
Подавляющее большинство инфузорий — гетеротрофы, хотя некоторые могут постоянно поддерживать в качестве симбионтов зеленые водоросли или хлоропласты, принадлежавшие съеденным простейшим, и получать необходимое питание за счет этих фотосинтетиков. В отношении питания представители Ciliophora также демонстрируют значительное разнообразие: среди них есть бактерио-, альго- и фунгиофа- ги, растительноядные, хищники, гистофаги и каннибалы (редко), причем встречаются как полифаги так и стено- и даже олигофаги. В то же время сами инфузории играют важную роль в пищевых цепях, служа добычей для многих мелких водных беспозвоночных. Обилие инфузорий как в планктоне, так и в бентосе приводит к тому, что им принадлежит значительная роль в круговороте азотистых соединений во всех типах водных (а часто и почвенных) экосистем (Laval-Pe- uto et al., 1986; Fenchel, 1987; Anderson, 1988).
Многочисленными исследованиями было показано, что инфузории могут служить индикатором качества воды и общего состояния водоемов, где они обитают (Curds, 1966; Bick, 1972; Antipa, 1977; Foissner, 1988). Некоторые виды этих простейших с успехом используются в качестве объектов при биотестировании (Инфузории в биотестировании, 1998).
Инфузории безусловно были в числе первых живых микроскопических организмов, обнаруженных и описанных Левенгуком (Dobell, 1932; Woodruff, 1945). С тех пор они прочно вошли в научный обиход, и помимо многих тысяч исследований, посвященных различным представителям этой группы простейших, некоторые инфузории стали излюбленными модельными объектами для решения самых разнообразных вопросов цитологии, молекулярной биологии, генетики, биохимии и физиологии.
История изучения
Начало изучения мира Protista неразрывно связано с изобретением и прогрессом микроскопической техники («мудрых стекол») в первой половине XVII в. Однако даже при наличии микроскопов, которые привлекали сначала внимание лишь узкого круга физиков, астрономов и философов (Соболь, 1949), должен был появиться исследователь, которого заинтересовал бы «невидимый мир живых существ». Прошло более пол столетия, прежде чем такой человек явился в лице дельфтского мануфактурщика и оптика-самоучки Антона ван Левенгука (1632—1723). Именно ему, несмотря на отсутствие какого-либо научного образования, принадлежат первые достоверные наблюдения живых инфузорий среди множества других микроскопических объектов, описанных им в письмах к Лондонскому Королевскому Обществу, начиная с 1674 г. (Dobell, 1932; Corliss, 1975, 2002). Левенгуку же принадлежит и первое изображение инфузории (очевидно, Nyctotheroides) из кишки лягушки (1683 г.). Эти поразительные для того времени наблюдения были получены с помощью простых микроскопов (по сути увеличительных луп) собственной конструкции, которые обладали, правда, значительным увеличением (до 300 раз). Вероятно, Левенгук, среди прочих технических новаций, первым добился в своих микроскопах и осветительного эффекта, напоминающего «темное поле»,(.'удя по подробным описаниям исследователя, видимо, он имел дело также с Carchesium, Chiliodonella, Coleps, Colpidium, Cyclidium, Dileptus, Kerona, Paramecium, Vorticella и некоторыми другими широко распространенными видами инфузорий (Corliss, 1975, 2002). При этом Левенгук отмечал не только морфологические детали наблюдаемых зверюшек («animalcules»), но и измерял их, а также описал их размножение, сократимость и, по-видимому, некоторые стадии конъюгации.
В это же время, правда, появилось еще несколько исследователей «невидимого мира». Например, в 1678 г. Христиан Гюйгенс (1629—1695) в письме к брату описал нескольких инфузорий (Dobell, 1932). Буонанни в 1691 г. впервые опубликовал изображение инфузории (по-видимому, Colpidium). Кинг двумя годами позднее изобразил нескольких простейших, включая Euplotes (Corliss, 1991).
В середине следующего, XVIII в., безусловно, доминировали воззрения отца современной таксономии и биологической номенклатуры Карла Линнея (1707—1778), который, будучи ботаником, мало интересовался водными «зверюшками» и не очень доверял микроскопу. Только в 12-м издании его «Systema Naturae» (1767) появилось 4 рода простейших, среди которых были персонифицированы Volvox и Vorticella. Все остальные Protozoa оказались включенными в два других рода с весьма красноречивыми названиями: Chaos и Furia. «Таинственные живые молекулы, разобраться в которых надлежит потомкам», — так охарактеризовал инфузорий Линней (Соболь, 1949). Однако еще в 1703 г. анонимный автор опубликовал очевидное изображение Paramecium (Woodruff, 1945; Wichterman, 1953). Эту же инфузорию исследовал Луи Жобло (1645—1723), который в 1718 г. первым описал у инфузорий ресничный покров, ядра и сократительные вакуоли. В 1752 г. Джон Хилл (1716—1775) в своей известной книге «История животных» дал «туфлеобразной» инфузории существующее поныне название — парамеция. Ему же принадлежат, среди многих других, названия Cyclidium и Enchelys. К сожалению, приоритет Хиллп не сохранен для истории, так как его описания были произведены за 6 лет до 1758 г., с января которого, согласно Международному коду зоологической номенклатуры, родовые названия признаются валидными, Следует упомянуть, что Абраам Трамбле (1710 1784), прославившийся изучением гидр, также в 1744....1748 гг, исследовал деление у «воронкообразных полипов» инфузорий рода Stentor и описал размножение Epistylis, Carchesium и Zoothamnium (Канаев, 1972; Corliss, 1991).
Вообще за XVIII в. достаточно много исследователей описывало и изображало в своих работах инфузорий, как и ряд других мелких одноклеточных и многоклеточных животных (жгутиконосцев, амеб, коловраток, личинок трематод), считавшихся тогда «наливочными животными» (infusoria) (см. Куторга, 1839). С. С. Куторга (1805—1861) — один из первых русских исследователей инфузорий писал: «это открытие (инфузорий) привело в движение всех исследователей, и всякий, кто имел микроскоп, спешил полюбоваться неутомимым движением этого незримого мира» (Куторга, 1839). Среди таких ученых были Бейкер, описавший в 1753 г. Lacrymaria, и Вризберг, который впервые (1765) использовал термин «инфузории» для простейших. В 1769 г. Эллис экспериментально вызвал экструзию трихоцист у Paramecium, а Гванцати впервые в 1796 г. описал цисты у инфузории, по-види- мому принадлежавшей к роду Amphileptus (Corliss, 1991).
Несомненно, особого внимания заслуживает научная деятельность Отто Фридриха Мюллера (1730— 1784) — знаменитого датского зоолога и, очевидно, первого систематика инфузорий. В его выдающихся монографиях «Vermium terrestrium et fluviatilium seu animaliun infusoriorum, helminthicorum et testaceorum» (1773) и «Animalcula infusoria fluviatilia et marina» (1786) приведено около 300 описаний видов различных протистов, а также бактерий и мелких многоклеточных, значительная часть которых принадлежит к морским и пресноводным инфузориям. Им были описаны инфузории из морского песка, скутикоцилиати- ды, гимностоматиды, тинтиниды, олиготрихи, суктории, колониальная Ophrydinium, имеющая домик морская Folliculina, а также приведены некоторые замечания по их экологии и физиологии. Названные в соответствии с биномиальной номенклатурной системой Линнея, многие инфузории до сих пор сохраняют в своем названии буквы О. F. М. Существенной заслугой Мюллера была правильная интерпретация феномена конъюгации инфузорий как сексуального процесса. Потребовалось, однако, почти 100 лет, чтобы протозоологи наконец приняли эту точку зрения.
К концу XVIII в. относится и первая работа, выполненная в области протозоологии русским автором. Зоолого-физиологическая диссертация М. М. Терехов- ского (1740—1796) «О Chaos infusorium Линнея», написанная в 1775 г., была защищена в Страсбургском университете (Соболь, 1949). Ее автор путем длительных, последовательных экспериментов с различными «инфузорными» настоями в 88 параграфах своей диссертации доказал, что инфузории «как все животные происходят путем размножения от предшествующих родителей». Таким образом, Тереховский еще в 1775 г. опроверг распространенную идею о самозарождении инфузорий (основные работы, выполненные на эту тему Спалланцани (1729—1799), появились в 1765 и 1776 гг.) и неопровержимо доказал их животную природу — вопросы, волновавшие ученый мир и в первой половине XIX в. «Пусть те, кому дано исследовать природу глубже, определят, относятся ли эти водные анималькули к червям или насекомым, или к другому классу животных — писал в заключении к своей работе Тереховский, — пусть они также, следуя Хиллу и Мюллеру, точнее определят их роды и виды» (Соболь, 1949). Этот завет был выполнен исследователями инфузорий следующего, XIX в.
XIX в. принес небывалый рост интереса к Protista, который реализовался прежде всего по двум направлениям. Продолжение описания новых видов протистов и уточнение деталей их морфологии, очевидно, было связано с расширением географии исследований и дальнейшим совершенствованием микроскопической техники: появление сложного объектива Шевалье, микроскопа Листера и коррекционного окуляра Росса (Карпов, 2001). Первые попытки классификации простейших на основе характеристик их групп (направление, положившее начало систематики Protozoa) оказались возможными лишь по достижении «критической массы» видовых описаний и с началом их сравнительного таксономического изучения (Corliss, 1991).
После О. Ф. Мюллера первым крупным исследователем и систематиком инфузорий был, несомненно, начавший свои работы в конце первой четверти XIX в., X. Г. Эренберг (1795—1876). После выпуска им фундаментальной монографии «Infusionsthierchen als Vol- lkommene Organismen» (Ehrenberg, 1838), в которой были приведены описания около 350 видов инфузорий (в тогдашнем понимании состава этой группы), снабженные множеством прекрасных иллюстраций, и развита «полигастрическая» теория их организации, Эренберг стал на десятилетие непререкаемым авторитетом в протозоологии. Собственно, как отдельная дисциплина в пределах зоологии она появилась именно благодаря его трудам. В то же время его теория инфузорий как совершенных организмов (обладающих всеми особенностями многоклеточных, но в миниатюре) безусловно послужила тормозом на пути развития учения о Protozoa — это название было введено в науку Гольдфусом (1782—1848) в 1817 г. (Goldfuss, 1817). Созданная накануне опубликования (1838— 1839) основ классической клеточной теории Шванна (1810—1882) и Шлейдена (1804—1881), теория Эренберга отражала «житейский» взгляд на возможности функционирования любого живого организма. Наличие в составе тогдашних инфузорий церкарий и коловраток дало дополнительный повод искать сходной организации у собственно инфузорий, и, конечно, она была обнаружена. Эренберг «открыл» у наливочных животных пищеварительный тракт (включая зубы и различные железы), мускулатуру, половые органы, глаза и зачатки нервной системы.
Система инфузорий в начале XIX в. в основном представляла собой разные модификации взглядов Ламарка (1744—1829) и Кювье (1773—1838), которые сами не занимались инфузориями и использовали для своих систем материал О. Ф. Мюллера. Так, Ламарк в 1815г. поместил инфузорий в два первых класса своей системы (не имеющие никакой организации или сходные с полипами). Кювье (1817) составил из инфузорий отдельный класс, разделив его на два отряда: Rotiferes (коловратки) и Infusoires homogenes (куда попали все простейшие). В книге Швейгера, выпущенной в 1820 г., мюллеровские инфузории попали в класс зоофитов, разбитый на два отряда: состоящие из «простого вещества» (инфузории) и «разновещественные» (полипы) (Куторга, 1839).
По Эренбергу, инфузории (Polygastrica) также должны были причисляться к зоофитам или лучистым, где составляли два класса: бескишечных (амебы, жгутиконосцы, диатомовые) и кишечных (собственно инфузории и некоторые солнечники). В пределах последней группы он выделил 4 отдела, положив в их основу лишь два признака: положение рта и порошицы (цито- прокта), что привело к крайне искусственной системе. Русский последователь Эренберга С. С. Куторга, составивший на основе трудов немецкого автора, с некоторыми собственными добавлениями, книгу «Естественная история наливочных животных» (Куторга, 1839), тем не менее, поместил инфузорий в класс членистых животных. В пределах этого класса он полагал их наиболее близкими с Entozoa (внутренностным или глистам). Отдел наливочных был разделен Куторгой на два отряда. К первому — дрожалкам (Vibratoria) — были причислены бескишечные (амебы и жгутиконосцы), а также кишечные (инфузории). Ко второму — коловратки (Rotatoria). Этой же схемы придерживался для инфузорий и другой российский ученый П. Ф. Го- рянинов (1796—1865), который, однако, поместил этих простейших в состав зоофитов (Горянинов, 1837).
При неправильной интерпретации строения инфузорий рисунки Эренберга поражают своей точностью, а его исследования географического распространения наливочных животных в Северной Африке, Аравии, Германии, Европейской и Азиатской России безусловно надолго стали главным источником информации по зоогеографии простейших. Таким образом, участвуя в путешествии по России, предпринятом А. Гумбольтом (1769—1859) в 1829 г., Эренберг стал первым исследователем инфузорий на бескрайних просторах нашего Отечества (от Санкт-Петербурга до Барнаула).
Младшим современником Эренберга был Феликс Дюжарден (1801—1860), который, помимо значительных достижений в исследовании амеб (он, собственно, предложил термин Rhizopoda), занимался и инфузориями и первым «восстал» против теории Эренберга (Dujardin, 1841). Однако его система деления инфузорий на симметричных (Coleps) и асимметричных (все прочие простейшие) оказалась не менее искусственной (Шевяков, 1896). С появлением деления животных на многоклеточных и одноклеточных (Siebold, 1845) и четкого определения понятия клетка (X. Моль, Ф. Лейдиг и М. Шульце) теория Эренберга к 60-м гг. XIX в. была «сдана в архив». Фактически к этому времени одноклеточные уже были разделены на два класса: Rhizopoda и Infusoria (Siebold, 1845), и позднее, говоря об инфузориях, исследователи имели в виду именно Ciliata — одноклеточных, снабженных мерцательными ресничками (Perty, 1852). Тогда же начал свои исследования инфузорий Э. Бальбиани, которому мы обязаны правильной оценкой феномена конъюгации у этих простейших (1861).
Значительный шаг вперед в изучении инфузорий был сделан в 50-е гг. XIX в. Клапаредом (1832—1871) и Лахманом. Эти ученые на основании собственных продолжительных и тщательных наблюдений издали в 1858—1861 гг. двухтомный труд «Les Infusoires et les Rhizopodes» (Claparede, Lachmann, 1858—1861), где выделили Suctoria в отдельную группу, а остальных Ciliata разбили на 10 семейств. Почти одновременно (в 1859 г.) появилась в печати и система Ф. Штейна (1818—1885), сыгравшая кардинальную роль в развитии цилиатологии позапрошлого, а, отчасти, и прошлого (XX) вв.
Штейн положил в основу классификации ресничный покров и на основании распределения ресничек по телу инфузорий разбил всех Ciliata на 4 отряда: Holo- tricha, l letcrotricha, Hypotricha и Peritricha. Flagellata и Suctoria также получили в его системе ранг отряда. Любопытно, что в одной из первых макросистем живых организмов Геккеля (1834—1919), где одноклеточные были уже объединены в царство Protista (1866), инфузории оказались включенными им на приведенной схеме в группу Articulata (т. е. высокоорганизованных многоклеточных!). Видимо, взгляды Эренберга, от которых он не отказался до конца жизни, еще находили себе сторонников.
Штейн, занимаясь исключительно инфузориями на протяжении нескольких десятков лет, описал много новых родов и видов, так что его система включала уже 23 семейства и 107 родов. В результате тщательного изучения представителей гипотрих и гетеротрих Штейном были выпущены две монографии, посвященные соответствующим отрядам (Stein, 1859, 1867), однако его блестящая серия осталась незавершенной. Дальнейшее развитие исследований инфузорий оказалось связанным сразу с несколькими учеными: это были О. Бючли (1848—1920) и Р. Гертвиг (1850— 1937) в Германии, Э. Мопа (1842—1916) во Франции и В. Кент (1845—1908) в Англии. Все они родились в одно десятилетие с 1842 по 1850 г. и во многом направление их работы совпадало, хотя след в науке они оставили не одинаковый.
О. Бючли — «архитектор протозоологии», как его назвал Добель (Dobell, 1951), и «гигант среди гигантов», по определению Корлиса (Corliss, 1978), начал изучать инфузорий с 1871 г. и, несмотря на то что он был зоологом широкого профиля, конечно, больше всего сделал именно в протозоологии. Надо сказать, что, по-видимому, интересом к инфузориям и фундаментальными знаниями в этой области Бючли был во многом обязан самообразованию: почти 4 года (1871, 1873—1876) он работал самостоятельно в собственной (частной) лаборатории в родном Франкфурте (Новиков, 1922; Fokin, 2004b). Уже в 1876 г. Бючли публикует работу, где впервые после домыслов Эренберга, Дюжардена и Клапареда были правильно поняты и описаны события, происходящие с ядерным аппаратом инфузорий при конъюгации. Помимо полового процесса инфузорий Бючли исследовал физиологию и морфогенез Ciliata, их жизненные циклы, цитологию. Последняя область исследований привела ученого к анализу строения протоплазмы, при моделировании которого Нючли применил, в том числе, и свои знания о коллоидной и физической химии. Его теория ячеистой структуры цитоплазмы, созданная в 90-х гг. XIX в, была широко распространена еще в первой четверти XX в, (Гартмин, 1929)
Нельзя не упомянуть, что школу Бючли как протозоолога прошло немало молодых людей, в разное время учившихся и риботинших II Гейдельберге, где Бючли был профессором зоологии 42 года. Среди них такие известные немецкие и русские протистологи и клеточные биологи, как Ф. Блохмапн, Р. Лаутерборн, М. Гартманн, К. Гамбургер, В, Т, Шевяков, С. И. Метальников, Н. К. Кольцов, С. В. Аперснцсв и А. С. Щепотьев (Fokin, 2004b).
Вершиной творчества Бючли-протистолога было написание трехтомного труда Protozoa (1880—1889), где третий том (Biitschli, 1887—1889) был почти целиком (953 стр.!) посвящен инфузориям. Там Бючли предложил свою классификацию класса Ciliata. Класс был разбит на два подкласса (Ciliata и Suctoria). Подкласс ресничных подразделялся на два отряда сообразно устройству рта и глотки (Gymnostomata и Trichoso- tomata), и последний по наличию адорального ряда ресничек был разделен на две группы в ранге подотряда: Aspirotricha и Spirotricha. Спиротрихи были в свою очередь разделены на четыре группы (следуя системе Штейна): Heterotricha, Oligotricha, Hypotricha и Peritricha. Все Ciliata включали 25 семейств, содержащих 142 рода. На тот момент это была самая совершенная система, в основе которой лежал комплекс признаков, а отряды и семейства представляли собой достаточно естественные группировки (Шевяков, 1896).
Э. Мопа не был профессиональным биологом, однако след, оставленный им в цилиатологии, также весьма значителен. В 80-е гг. XIX в. он опубликовал серию замечательных работ, посвященных кариологии, половому процессу и размножению у инфузорий (Maupas, 1883, 1886, 1888).
Р. Гертвиг был известен не только как протистолог, но и как экспериментальный эмбриолог. Очевидно, от своего учителя Э. Геккеля он унаследовал интерес к простейшим, среди которых исследовал прежде всего радиолярий, инфузорий и солнечников. В области цилиатологии он заслуживает памяти как автор очень глубокой работы, посвященной конъюгации инфузорий (Hertwig, 1889). Так же как и Бючли, Гертвиг был прекрасным педагогом, и среди его учеников было немало видных протистологов.
В. С. Кент был разносторонним зоологом, и инфузории являлись лишь одной из многих групп беспозвоночных и позвоночных животных, над которыми работал этот английский ученый (Esteban et al., 2002). В цилиатологии он оставил след прежде всего изданием фундаментальной серии (3 тома) «А Manual of the Infusoria» (Kent, 1880—1882) и описанием значительного числа видов среди перитрих и сукторий. Последние были выделены Кентом в отдельную группу Tentaculi- fera (Corliss, 1978).
Приблизительно в это же время исследования инфузорий начались и в Новом Свете. Наиболее заметной фигурой среди цилиатологов США был в конце XIX в. А. С. Стокс. Он выпустил солидную сводку по фауне инфузорий Соединенных Штатов, а также написал учебные пособия по изучению микроскопических объектов (Stokes, 1888, 1894, 1896).
В России вторая половина XIX в. была ознаменована появлением исследователя, вполне сравнимого по широте взглядов и глубине мысли с лучшими представителями Западноевропейской школы. Это был выпускник Петербургского университета, а потом, к несчастью лишь недолгое время (1854—1861), и его профессор, Л. С. Ценковский (1822—1887) — выдающийся русский ученый (польского происхождения), один из основателей отечественной протистологии и микробиологии (Райков, 1959; Kuznicki, 1982; Fokin, 2001b). Его рассуждение «О строении простейших живых организмов», представленное для получения доцентуры в 1847 г. (Райков, 1959), и докторская диссертация «О низших водорослях и инфузориях» (Ценковский, 1856) включали исследования различных протистов, в том числе и инфузорий (Enchelys, Stylonychia и Vorticella), и убедительно показали, что простейшие состоят из одной клетки и соответственно не имеют никаких органов, приписываемых им Эренбергом. Ученый, по сути введший в российскую практику университетского преподавания микроскоп, Ценковский открыл и описал несколько десятков различных протистов и проследил для многих из них жизненные циклы. Одним из первых он исследовал процессы цистообразования у Ciliata и экспериментально доказал невозможность самозарождения у простейших (Ценковский, 1859). В своих публичных лекциях Ценковский проводил мысль о связи одноклеточных и многоклеточных животных и первым обратил внимание на явление симбиоза у низших организмов (Райков, 1959; Fokin, 2001b). Обстоятельства жизни не позволили ему создать реальную протистологическую научную школу. Во второй части своей научной карьеры (в Одессе и особенно в Харькове) Л. С. Ценковский переключился на микробиологические исследования (Метелкин, 1950). Однако его протистологические работы были продолжены рядом учеников и последователей (А. С. Фаминцын, А. О. Вер- жесниовский, М. С. Воронин и К. С. Мережковский). В том числе, возможно под влиянием работ Ценковско- го, И. И. Мечников (1845—1916) начал свою научную карьеру именно с изучения инфузорий, опубликовав несколько своих наблюдений над этими простейшими (Мечников, 1864а, б, 1865; Metchnikoff, 1864).
Среди перечисленных ученых К. С. Мережковский (1855—1921) в первой части своей научной карьеры (1877—1886) наиболее целенаправленно занимался именно инфузориями. Ему принадлежат первые (после серии работ Э. И. Эйхвальда 1844—1860 гг.) фаунистические сводки по простейшим России: «Этюды над простейшими Севера России» (1878), «Материалы для фауны инфузорий Черного моря» (1880) и «О некоторых новых или малоизвестных инфузориях» (1881).
Достойно упоминания, что среди русских исследователей инфузорий последней четверти XIX в. было две женщины, случай по тем временам чрезвычайно редкий. Это были Ю. И. Андрусова (1863—1942) и С. М. Переяславцева (1849—1903), которые также занимались изучением фауны инфузорий Черного моря (Андрусова, 1886; Переяславцева, 1886). Известный физиолог растений, ученик Л. С. Ценковского, А. С. Фаминцын (1843—1918) также неоднократно обращался к исследованию простейших, и преимущественно инфузорий (Фаминцын, 1889, 1890).
В конце XIX в. активные исследования инфузорий проводились также в Варшавском, Киевском и Новороссийском университетах. В первом долгое время (1864—1889) работал известный польский протисто- лог А. О. Вержесниовский (1837—1892), хотя пик ци- лиатологических работ там пришелся на время заведования зоотомическим кабинетом его преемника, проф. П. И. Митрофанова (1857—1925). Помимо фаунистических работ (Эйсмонд, 1890) существенное внимание исследователи в Варшаве уделяли вопросам цитологии инфузорий (Пржесмыцкий, 1894; Сосновский, 1897; Кудельский, 1898; Мавродиади, 1913).
Большую работу по морфологии и биологии щупальцевых инфузорий опубликовал киевлянин Н. А. Кеп- пен (1888), а П. П. Бучинский (Одесса) занимался изучением протистофауны лиманов (Бучинский, 1895). Один из учеников Мечникова по Новороссийскому университету В. М. Хавкин (1860—1930), ставший впоследствии известным бактериологом, положил начало изучению бактериальных эндобионтов инфузорий (Hafkine, 1890).
С осени 1894 г. цилиатология в России получила значительное подкрепление — в Петербург из Германии вернулся В. Т. Шевяков (1859—1930). Окончив Гейдельбергский университет, где он учился у О. Бюч- ли (перед этим 4 года он был студентом-вольнослушателем у Н. П. Вагнера в С.-Петербургском университете), Шевяков еще в Германии начал успешные исследования инфузорий (Fokin, 2000, 2004b; Фокин, 2001). Сразу по окончании университета он совершил кругосветное путешествие (1889—1890). Главной целью этой десятимесячной поездки было исследование географического распространения пресноводных простейших, и прежде всего Ciliata. Написанная по материалам поездки монография (Schewiakoff, 1893), была через 3 года удостоена в России Брантдовской академической премии. По возвращении на родину Владимир Тимофеевич защитил магистерскую диссертацию «К биологии простейших», опубликованную в виде пространной статьи, две (из трех) глав которой были посвящены морфологии и распространению инфузорий (Шевяков, 1894).
Через 2 года В. Т. Шевяков представляет в качестве докторской диссертации работу «Организация и систематика Infusoria Aspirotricha (Holotricha auctorum)» (1896), которая блестяще завершила серию монографий, начатую Ф. Штейном. Плод десятилетних наблюдений и размышлений ученого (408 страниц текста и 7 больших таблиц превосходных рисунков) включил сравнительно-анатомическое описание морфологии инфузорий, анализ ряда биологических и физиологических особенностей группы, их географическое распространение и подробное обсуждение системы и филогенетических связей холотрих.
Классификация инфузорий, предложенная Шевяковым, несколько отличалась от системы Бючли. Так, Holotricha представлялась ему более естественной группой, не заслуживающей разделения на Gymnosto- mata и Trichostomata в ранге отрядов. Все Ciliata были разделены Шевяковым на 2 отряда: Spirotricha и Aspirotricha. Их состав, естественно, изменился. Отряд Aspirotricha, рассматриваемый Шевяковым как более примитивный, включил в себя Holotricha Штейна, с некоторыми добавлениями. На основе устройства рта и глотки у представителей этого отряда он был разбит на 3 подотряда: Gymnostomata, Trichostomata и Astomata. Всего отряд Aspirotricha был разделен Шевяковым на 19 семейств, содержащих 80 родов и 181 вид. Несколько таксономических групп, выделенных Шевяковым: Astomata, Prostomata, Pleurostomata (Шевяков, 1896), сохранились в системе Ciliophora до наших дней (Lynn, Small, 2000).
С момента появления Шевякова Петербургский университет становится крупнейшим протозоологиче- ским центром России. Многие ученики Владимира Тимофссвича отдали дань научной тематике своего профессора. Среди них такие известные ученые XX в., как С. И. Метальников, М. Н. Римский-Корсаков, Л. К. Линько, С. В. Аверинцев, В. А. Догель, И. Н. Филиньсн и И. К. Дембовский.
Начало XX в. принесло значительное расширение гос гана и географии сообщества протозоологов-цили- п голого». Помимо уже названных ученых на научное поприще вступили их ученики и младшие коллеги. Большое значение для консолидации научных сил, обмена свежей информацией и привлечения новых исследователей имел первый международный прото- зоологический журнал «Archiv fur Protistenkunde» (1902), организованный Ф. Шаудином (1871—1906), а также появление первых учебников по протозоологии (Calkins, 1901; Doflein, 1901). Впоследствии в России также был создан протистологический журнал («Русский архив протистологии»), игравший аналогичную роль, но, к сожалению, просуществовавший лишь 8 лет (1922—1929).
Г. Н. Калкинс (1869—1943) был первым профессором, начавшим читать специальный курс «Протозоология» и возглавившим одноименную кафедру. Сам он занимался по преимуществу физиологией и клеточными циклами простейших и работал прежде всего с парамециями. В 1912 г. его учебник был переведен на русский язык (Калкинс, 1912) московским зоологом В. С. Елпатьевским.
Ф. Дофляйн (1873—1924), ученик Р. Гертвига, имел весьма широкий круг научных интересов, в том числе занимался и эпибионтными инфузориями Chonotricha (Янковский, 1973). Именно ему принадлежит выделение инфузорий в отдельный тип Ciliophora (Doflein, 1901). Другой немецкий протозоолог, работавший и у Бючли, и у Гертвига, — М. Гартманн (1876—1962) выпустил «Практикум по протозоологии» (Hartmann, 1907). Гартманн, занимавшийся вначале систематикой и сриниительной морфологией различных простейших, и том числе и инфузорий, впоследствии сосредоточился и» теоретических вопросах. Исследование полового процесса у простейших и анализ организации их ядер интересовали Гартманна на протяжении всей ого долгой ппучной жизни (Hartmann, 1952, 1955). Это ииприилемио активно развивал и один из ближайших сотрудников Гиргманна — К. Беляр (1895—1929), чья первни книги о ядре простейших (Belar, 1926) обещала ему блестящее научное будущее, к несчастью трагически несостоявшееся. Влияние упомянутых ученых пи развитие цитопатологии трудно переоценить. То же самое следует сказать и о самой удивительной фигуре в систематике И фаунистике Ciliophora первой трети XX в. А. Кило (1877 -1946).
Будучи учителем гимназии и занимаясь изучением инфузорий н качестве хобби (Foissner, Wenzel, 2004), Каль опубликовал свою норную работу в 49 лет (Kahl, 1926). За 9 лет в серии «Животный мир Германии» и «Животные Северного и Западного морей» появилось свыше 1000 страниц видовых описаний, ключей для определения и рисунков, выполненных им без всяких помощников и отражавших практически всю совокупность данных по известным к тому времени свободно- живущим и эктокоммсисальпым инфузориям (Kahl, 1930—1935, 1933,1934, 1934а). Последняя крупная работа А. Каля, опубликованная в 1943 г., была обнаружена совсем недавно (Kahl, 1943; Foissner, Wenzel, 2004). Им была проведена ревизия системы Ciliophora и предложена ее новая версия, вобравшая в себя достижения Штейна, Бючли, Шевякова и Шаттона — неимоверная работа подвижника-одиночки!
Во Франции первая четверть XX в. также ознаменовалась появлением выдающихся исследователей, особенно много работавших в области изучения инфузорий: Э. Шаттона (1883—1947) и Э. Фаре-Фремье (1883—1971). Оба они широко использовали в своих исследованиях разработанный Шаттоном метод импрегнации кортекса инфузорий серебром (Chatton, Lwoff, 1930). Шаттону, который изучал прежде всего морские и симбиотические формы, принадлежит открытие правила десмодексии, установление основных типов деления протистов и выявление автономности процесса умножения кинетосом у инфузорий. Он впервые отчетливо осознал эволюционное расстояние между бактериями и другими организмами, предложив термины про- и эукариоты. Ряд работ по цилиатологии были выполнены Шаттоном совместно с будущим нобелевским лауреатом А. Львовым (1902—1994), потомком выходцев из России, который особенно в начале своей научной карьеры много занимался именно инфузориями (Chatton, Lwoff, 1922, 1935, 1939) и сохранил интерес к этой тематике до конца своей долгой жизни (Soyer-Gobillard, Schrevel, 2003).
Фаре-Фремье, работавший по преимуществу с пресноводными свободноживущими простейшими, использовал обнаруженные коллегой особенности кортикального морфогенеза инфузорий для анализа эволюции и филогении Ciliophora. Он уделял особенное внимание процессам стоматогенеза в различных группах инфузорий, что позволило ему позднее создать новый вариант системы этого типа (Faure-Fremiet, 1950а).
Надо отметить, что, помимо традиционно «сильной» немецкой и французской школ цилиатологов, с начала XX в. сформировалась замечательная плеяда американских исследователей инфузорий. Параллельно с Г. Калкинсом начал работать в университете штата Иллинойс С. А. Кофойд (1865—1947), занимавшийся прежде всего паразитическими простейшими и эндокомменсалами. Он исследовал также инфузорий из рубца домашних жвачных и Entodiniomorpha слонов и занимался тинтинидами и «нейромоторным» аппаратом инфузорий и жгутиконосцев (Corliss, 1978). Другим видным протозоологом этого же поколения был Г. С. Дженнингс (1868—1947). Специалист в области поведения и физиологии простейших, он работал преимущественно на инфузориях. Дженнингс одним из первых начал исследования по генетике Ciliophora (Jennings, 1929) и стал использовать в качестве объекта Р. bursaria (Jennings, 1939). Первый студент Калкинса Л. Л. Вудруф (1879—1947) также работал исключительно на инфузориях, занимаясь в основном вопросами их цитологии и жизненных циклов. Кроме того, он опубликовал ряд работ по истории зоологии, и в том числе по истории исследования парамеций (Woodruff, 1926, 1938, 1945).
В России начало XX в. ознаменовалось дальнейшим развитием Санкт-Петербургской школы протозоологов. Среди ее представителей наиболее выделялся В. А. Догель (1882-1955), ставший признанным лидером отечественной протозоологии в советское время. В области цилиатологии он работал исключительно над представителями семейства Ophryoscolecidae и изучил эту группу досконально (Догель, 1923, 1925а, 1925в, 1927). Его многочисленные ученики также исследовали инфузорий (Тассовский, 1916, 1918; Раммельмейер, 1925; Полянский, 1928; Стрелков, 1928; Фурсенко, 1928; Богданович, 1930; Жинкин, 1930; Хейсин, 1930).
Интересные работы по цитологии, фаунистике и физиологии некоторых Ciliophora опубликовали в первой четверти XX в. С. И. Метальников (1907, 1911; Метальников, Галаджиев, 1916), Н. К. Кольцов (1911), Б. А. Сварчевский (1912, 1928) и Л. Л. Россолимо (1916), Л. Л. Россолимо и Т. А. Перцева (1929). Обширные эколого-фаунистические исследования инфузорий развернула на Байкале Н. С. Гаевская (1927, 1933).
Середина и конец XX в. в истории цилиатологии настолько богаты разнообразными событиями, а круг исследователей и направления их работы в это время настолько расширились, что этот период требует специального, большого обзора.
Общее число работ, выполненных на различных Ciliophora, не поддается учету. Достаточно сказать, что только с использованием Paramecium за 250 лет (преимущественно в XX в.) было выполнено свыше 5 тыс. исследований. Несмотря на то что работы, сделанные на инфузориях в XVIII—XIX вв., частью не потеряли своего научного значения и, конечно, представляют значительный исторический интерес, я буду ссылаться в настоящем разделе лишь на исследования XX в., как на наиболее соответствующие современному уровню протистологической науки.
Воистину всемирную известность снискали такие представители инфузорий, как Paramecium и Tetrahy- тепа. Результатом длительного и глубокого изучения некоторых Ciliophora и неослабного интереса к этим простейшим со стороны как протистологов, так и ученых других специальностей можно считать целую серию публикаций, посвященных монографическому описанию ряда инфузорий: Paramecium (Kalmus, 1931; Wichterman, 1953, 1986; Beale, 1954; Jurand, Selman, 1969; Wagtendonk, 1974; Janisch, 1987; Gortz, 1988); Tetrahymena (Elliot, 1973; Asai, Forney, 2000); Blepha- risma (Giese, 1973); Stentor (Tartar, 1961); Ophryoglena (Canella, Rochi-Canella, 1976); Colpodea (Foissner, 1993).
Различные аспекты систематики, биологии и молекулярной биологии инфузорий нашли свое отражение в фундаментальных книгах Калкинса (Calkins, Sommers, 1941), Львова (Lwoff, 1950), Корлисса (Corliss, 1961, 1979), Нэнни (Nanney, 1980), а также в крупных сводках и коллективных монографиях (Серавин, 1967; Райков, 1978; Осипов, 1981; Small, Lynn, 1985; Лукашенко, Рыбакова, 1986; Gall, 1986; Frankel, 1989; Lynn, Corliss, 1991; Hausmann, Bradbury, 1996; Hausmann, Hiilsmann, 1996; Hiwatashi, 1999; Lynn, Small, 2000; Hausmann et al., 2003).
Значительное число монографий и руководств было посвящено фауне инфузорий отдельных мировых регионов, стран, морей, озер и рек, а также инфузориям определенных экологических систем и систематических групп (Gajewskaja, 1933; Bhatia, 1936; Чорик, 1968; Dragesco, 1965, 1966, 1970; Янковский, 1973; Мамаева, 1979; Curds, 1969, 1982; Ogimoto, Imai, 1981; Curds etal., 1983; Агамалиев, 1983; Maeda, Carey, 1985; Maeda, 1986; Dragesco, Dragesco-Kemeis, 1986; Локоть, 1987; Matthes et al., 1988; Бойкова, 1989; Carey, 1992; Foissner, 1994; Petz et al., 1995; Довгаль, 1996; Корнилова, 2004). Ряд этих изданий снабжены лишь списками обнаруженных видов (Patterson et al., 1989; Dini et al., 1995; Жариков, 1996; Жуков и др., 1998; Yingzhi, Yun Fen, 2000; Fokin, 2001a), в то время как другие включают значительное число иллюстраций и содержат исчерпывающие данные по морфологии, систематике и экологии большого числа представителей исследованных групп Ciliophora (Янковский, 1973; Foissner et al., 1991, 1992, 1994, 1995, 1999, 2002).
В некоторых монографиях и руководствах имеется весьма важный материал по методам выделения живых культур инфузорий из природы, их поддержания в лабораторных условиях и основным приемам изучения и идентификации Ciliophora (Иванов и др., 1981; Finlay et al., 1988; Lee, Soldo, 1992; Foissner, 1993; Жариков, 1996; Алекперов и др., 1996).
Несколько международных и национальных коллекций живых культур микроорганизмов содержат в том числе и некоторый материал по Ciliophora (АТСС, ССМР, США; SCCAP, Дания; ССМ, Чешская Республика; ССАР, Великобритания). Существуют значительные коллекции ряда видов инфузорий и в России (например, в БиНИИ СПбГУ, Санкт-Петербург). Хотя, как правило, исследователи ограничиваются поддержанием лишь непосредственных объектов своей работы, а многие экологи, гидробиологи и исследователи протистофауны вообще никогда не создавали живых коллекций обнаруженных простейших.
Результаты исследований, посвященных Ciliophora, печатаются в ряде специальных периодических изданий: Protist (с 1902 по 1997 — Archiv fur Protisten- kunde), Journal of Eukaryotic Microbiology (c 1954 no 1988 — Journal of Protozoology), European Journal of Protistology (c 1965 no 1986 — Protistologica), ActaPro- tozoologica (c 1963), Japan Journal of Protozoology (c 1967), Protistology (c 1999) и в многочисленных международных и отечественных журналах, тематически связанных с изучением простейших.
Конечно, инфузории — одна из ключевых групп царства Protista, постоянно упоминаются в многочисленных учебниках, учебных пособиях и сводках как по протистологии и протозоологии, так и по зоологии беспозвоночных, экологии и эволюции (Hyman, 1940; Догель, 1951; Grasse, 1952—1953; Doflein, Reichenaw, 1949—1953; Mackinnon, Hawes, 1961; Догель и др., 1962; Pitelka, 1963; Raabe, 1964; Lipsi, 1965; Райков, 1967, 1978; Chen, 1967—1972; Kudo, 1971; Grell, 1973; Fanner, 1980; Puytorac et al., 1987; Puytorac, 1994; Хаусман, 1988; Sleigh, 1989; Карпов, 1990; Margulis et al., 1990; Mehlhorn, Ruthmann, 1992; Foissner, 1994; Coombs et al., 1998; Карпов, 2001; Корнилова, 2004).
Как видно из списка литературы, цитированной выше, число отечественных сводок по Ciliophora весьма мало, и в то же время значительный интерес к инфузориям, безусловно, требует появления таких работ. При этом следует отметить, что многие из перечисленных выше зарубежных изданий малодоступны даже в библиотеках столичных городов России. Таким образом, настоящий обзор призван, хотя бы отчасти, восполнить существующий дефицит русских источников но морфологии, биологии и систематике представителей одной из интереснейших групп протистов — инфузориям, а также дать представление об основных iTillinx истории развития цилиатологии в течение пер- ных 250 лет ее существования. Эта история, несмотря ни ряд публикаций (Calkins, 1901; Бродский, 1937; Corliss, 1978, 1991; Kuznicki, 1982, 2003; Entzeroth, 1994; Geus, 1994; Leadbeater and Green, 2000; Vickerman et al., 2000; Fokin, 2001b; Wolf and Hausmann, 2001; Корнилова, 2004), до сих пор слабо освещена в литературе.
Морфофункциональная организация
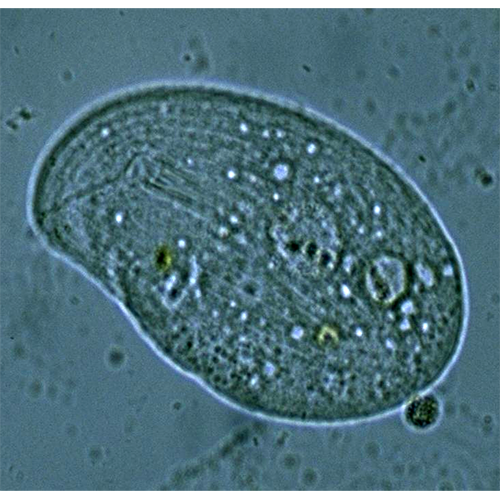
В водных экосистемах инфузории — наиболее заметные представители Protista, благодаря их сравнительно крупным размерам и подвижности. Как и все эукариотические клетки, они имеют стандартный набор клеточных органелл и структур, хотя некоторые особенности организации Ciliophora присущи только этим простейшим.Тело инфузорий состоит из двух основных ком- партментов: цитоплазмы и карио- или нуклеоплазмы. Первая отделяется от внешней среды особым наруж- ним слоем — кортексом, куда входит и типичная плазматическая мембрана, а вторая (от цитоплазмы) — ядерной оболочкой. Поскольку клетки Ciliophora имеют ядра двух типов (макро- и микронуклеусы), кариоплазма инфузорий представлена также двумя составляющими — макро- и микронуклеоплазмой, каждая из которых обычно отделена от цитоплазмы отдельной ядерной оболочкой — производной гладкого эндоплазматического ретикулума. Мелкий микронуклеус (или несколько микронуклеусов) содержит сравнительно сильно конденсированный хроматин, который часто рассматривают как неактивный, хотя, по-видимому, это не совсем так (Fokin, Ossipov, 1981; Ng, 1986; Фокин, 2002). Крупный единственный макронуклеус, иногда разделенный на связанные между собой или полностью автономные многочисленные фрагменты (Heterotrichia, Hypotrichia), содержит хроматин разной степени конденсации, активный на протяжении всего жизненного цикла клетки (Raikov, 1995, 1996).
Цитоплазматическая составляющая клетки инфузорий разделена на ряд субкомпартментов, связанных между собой. Кортекс клетки представлен поверхностным слоем толщиной 1—4 мкм, включающим пелликулу, которая песет наружную цилиатуру и инфраци- лиатуру. Иногда поверхностную зону цитоплазмы обозначают как эктоплазму. Граница между кортексом (эктоплазмой) и эндоплазмой у большинства инфузорий выражена нечетко, хотя у представителей некоторых групп (Litostomatea) между экто- и эндоплазмой залегает хорошо различимый филаментозный слой. В эндоплазме расположены ядра, пищеварительные и сократительные вакуоли, гладкий и шероховатый эндоплазматический ретикулюм, комплекс Гольджи, митохондрии (как правило) и некоторые другие органеллы и включения.
Организация клетки инфузорий, как и других протестов, может рассматриваться на разных ее уровнях: макромолекулярном, суборганелльном, органелльном, системно-органелльном и клеточно-организменном (Riedl, 1978; Lynn, Corliss, 1991), хотя не во всех случаях эти уровни одинаково хорошо выявляются. В общей характеристике организации клетки инфузорий, я буду оперировать в основном понятиями суборганелльного, органелльного и системно-органелльного уровней. Объяснение некоторых терминов дано в тексте и в специальном приложении. Развернутое толкование многочисленных специальных терминов и понятий, связанных с морфологией инфузорий, приведено в монографии Корлиса (Corliss, 1979), иллюстрированном глоссарии Protoctista (Margulis et al., 1993) и иллюстрированном гиде Protozoa (Lynn, Small, 2000).
КОРТЕКС
Особенности кортикальной организации инфузорий определяют собственно внешний вид клетки этих простейших. Кортекс несет барьерную функцию, отделяя внутреннюю среду организма от среды внешней. Он подразделяется на две составляющие: оральный и соматический, соотношения и особенности которых значительно варьируют в разных группах инфузорий.Плазматическая мембрана. Поверхность кортекса представлена плазматической (клеточной) мембраной толщиной около 10 нм со слоем гликокалекса, который может модифицироваться клеткой в ответ на изменение условий внешней среды. Эта мембрана имеет типичное трехслойное строение и несет значительное число внутримембранных частиц определенной топографии и архитектоники. Функции этих частиц пока не ясны. Гликокаликс, который состоит в основном из мукополисахаридов, также имеет характерную структуру и связан с коммуникативной системой клетки инфузорий. В нем расположены многочисленные рецепторные молекулы, что позволяет клетке-организму селективно абсорбировать из окружающей среды различные вещества. Иногда снаружи плазматической мембраны клетку инфузорий (особенно у некоторых гипотрих и олиготрих) покрывает еще одна мембраноподобная структура, — перилемма функция которой также неизвестна (Hausmann, Hiils- mann, 1996).
В совокупности со структурами, расположенными под плазматической мембраной: альвеолами, эпиплазмой и сопутствующими микротрубочками (общей толщиной менее 1 мкм), этот слой кортекса именуется пелликулой. В пределах типа, композиция пелликулы может варьировать. Даже в одной клетке в зависимости от зоны кортекса (несущей реснички или безресничной) пелликула может не совпадать по своей сложности. В целом архитектоника пелликулы у инфузорий также может различаться. У многих инфузорий она образует различные складки и гребни или, как у Paramecium и других пе- никулин, имеет вид гексогональных (иногда прямоугольных) ячеек, в центре которых расположены реснички, а в окружающих ячейку гребнях — трихоцисты.
Альвеолы. Альвеолы пелликулы инфузорий представляют собой окруженные мембраной уплощенные мешочки, образующие слой сразу под плазматической мембраной. Эти составляющие пелликулы характерны, кроме инфузорий, для Apicomplexa и Dinoflagellata. Однако даже среди инфузорий развитие этих структур может быть не одинаково: они не найдены у Karyorelictea, слабо представлены у некоторых Spirotrichea, Litostomatea и Phyllopharyngea (Chonotrichia) и в то же время достигают значительного развития и сложности организации у большинства представителей других классов Ciliopho- га. Топография и размеры альвеол не одинаковы у разных видов инфузорий, но сходны в клетках, принадлежащих к одному виду.
Альвеолы большинства инфузорий представляются пустыми при электронно-микроскопическом исследовании и их функции остаются пока не до конца ясными. Предполагается, что они могут быть связаны с регуляцией перемещения ионов, определяющей биение ресничек. Некоторые инфузории тем не менее содержат в альвеолах определенный материал. Так, клетки Coleps содержат в альвеолах карбонат кальция. Nassula и сходные формы класса Nassophorea имеют альвеолы, заполненные плотным веществом неясной природы, а Euplotes содержит в альвеолах плотный фиброзный материал, сходный по консистенции с расположенным в эпиплазме этих инфузорий.
Эпиплазма. Ниже альвеолярного слоя в кортексе инфузорий обычно представлен слой фиброзного или филаментозного материала, называемый эпиплазмой. Этот слой может быть выражен у разных инфузорий в неодинаковой степени, а иногда, возможно, отсутствует совсем. Отчасти степень выявления эпиплазмы зависит от характера подготовки материала для микроскопирования, поэтому говорить об отсутствии эпиплазмы в случаях, когда она не выявляется при электронно-микроскопическом исследовании, следует с известной осторожностью. По-видимому, эпиплазма играет важную роль в поддержании общей формы тела инфузорий, являясь частью цитоскелета. По крайней мере у мелких коль- подид, сохранение формы тела которых не требует дополнительных структур, она выражена слабо. В то же время крупные виды инфузорий, как правило, имеют сильно развитый слой эпиплазмы (Lynn, Corliss, 1991). В местах локализации цитофарингса, цитопрокта, пор сократительных вакуолей и экструсом эпиплазма обычно не выражена. Впрочем, эпиплазма может быть слабо представлена и в случае значительного развития других армирующих структур в пелликуле — микротрубочек или микрофиламентов.
Микротрубочки. Пелликулярные микротрубочки обычно достаточно многочисленны в кортексе инфузорий. Значительная их часть связана с кинетосомами, но как в несущих реснички частях кортекса, так и в лишенных ресничек областях поверхности инфузорий могут встречаться микротрубочки, не связанные с органеллами локомоции (Sundermann, Paulin, 1981; Lynn, Zimmerman, 1981). Пелликулярные микротрубочки иногда располагаются выше эпиплазмы, но в других случаях подстилают ее. Супраэпиплазматические микротрубочки характерны для представителей класса Oligohymenophorea (Tetrahymena, Colpidium, Colpoda, Conchophthirus), у которых они могут иметь вид тяжей между кинетами, образующих специфический слой в районе цитофаригса. Встречаются они и изолированно друг от друга (Allen, 1967; Antipa, 1971; Lynn, Zimmerman, 1981; Paulin, 1996). Субэпиплазматические микротрубочки наиболее характерны для участков тела инфузорий, лишённых цилиптуры, и соответственно такие образования нииболее часто встречаются у хинотрис, филофартеид и сукторий. У значительного числа инфузорий пелликулярные микротрубочки редки или их появление связано с определенной фазой клеточного цикла. Так, например, у Paramecium супра-магические микротрубочки появляются лишь в процессе клеточного деления, а в интерфазе исчезают (Sundararaman, Hanson, 1976).
Микрофиламенты. Роль кортикальных микрофиламентов, как уже было сказано, прежде всего формообразующая. Кроме того, они имеют большое значение при клеточном делении, составляя в его плоскости сократимое кольцо. Это образование, сжимаясь, осуществляет перешнуровку тела инфузории, в результате чего возникают две дочерние особи. безусловно, природа филаментов, обнаруженных в кортексе инфузорий, может быть различной, даже сени они участвуют в одних процессах. В некоторых случаях у инфузорий был выведен первичный актин (Cohen et al., 1980), но часто принцип работы сократимых элементов инфузорий отличается от актиномизионного типа. Так, спазмий, обнаруженный в спазмонемах стебельков представителей подкласса Peritrichia, обеспечивается чрезвычайно быстрое сокращение этих образований. У Striitur, Spirostomum и ряда других НеIeiolrichia за сокращение тела отвечают иные типы филаментов, образующих продольно ориентированные мионемы. Во всех перечисленных случаях сокращение этих клеточных элементов не связано с Л ГФ, по оказывается кальций-зависимым.
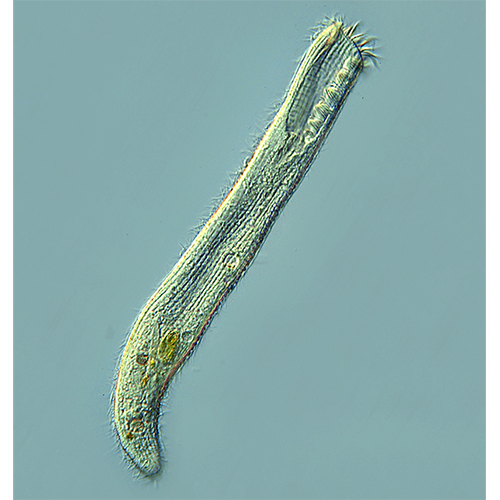
С соматическим отделом кортекса связано значительное число постоянных компонентов клетки инфузорий. Это прежде всего кинетиды, различного рода экструсомы и включения (пигментные гранулы), а также цитопрокт и поры сократительных вакуолей, если таковые вакуоли имеются. Соматическая часть кортекса инфузорий служит прежде всего для плавания, для прикрепления к субстрату, выделения защитных образований на поверхности клетки, а также для сенсорного взаимодействия с окружающей средой. Основной единицей соматической части кортекса является кинетида. Это комплексная структура, включающая базальное тело (кинетосому), три типа фибриллярных дериватов, которые с ней связаны, мембранное углубление (парасомальный сак) и собственно ресничку (Corliss, 1979; Lynn, Corliss, 1991; Paulin, 1996).
Реснички. Ресничка представляет собой окруженную мембраной совокупность микротрубочек (аксонему), включающую девять периферических дублетов (А- и Б-микро- трубочки) и один центральный (9+2). В то время как структурные модификации кинетосомы и аксонемы достаточно редки, переходная зона ресничек оказывается сравнительно пластичной. При совпадении основной ее функции (прикрепление реснички) к настоящему времени известно свыше 10 типов организации этой структуры у Protista, в том числе несколько для Ciliophora (Grain et al., 1988; Карпов, 1990, 2001; Фокин, 1994, 1995; Karpov, Fokin, 1995). В этой зоне реснички — участке кинетиды, расположенном в месте перехода из клетки во внешнюю среду, — выделяют систему поперечных пластинок, колец и иногда спиралей и «колес», а также аксосому с проксимальными концами центральной пары микротрубочек аксонемы. В переходной зоне реснички центральные микротрубочки заканчиваются, причем одна из них входит в аксосому (аксиальное зерно), а вторая оканчивается в аксонемном цилиндре слепо.
Морфологическое богатство структур, связанных с переходной зоной, и их консерватизм позволяют с успехом применять этот признак в таксономии и филогенетике групп ранга тип—класс (иногда подкласс). Организация переходной зоны ресничек у разных инфузорий закономерно различается в деталях. Так, у большинства представителей Peniculia (Oligohy- menophora) обнаружена одна из наиболее сложных (среди инфузорий) форм этого отдела ресничек, характерной особенностью которой служит не только наличие аксосомного кольца, но и присутствие концентрической структуры, напоминающей переходную спираль (Фокин, 1994; Karpov, Fokin, 1995). Эта структура располагается между промежуточной и аксосомной поперечными пластинками и состоит из четырех оборотов тонкой фибриллы, связанной с А-микротрубоч- ками. Достаточно сложные и характерные структуры переходной зоны ресничек отмечены также для Colpodea и Nassophorea (Фокин, 1995).
Некоторые детали строения мембраны ресничек (количество и топография межмембранных частиц) дают известный материал для сравнительного анализа инфузорий (Bardele, 1981). При использовании техники замораживания—скалывания на протоплазматической части мембраны ресничек отчетливо заметны группы межмембранных частиц различной архитектоники: «кружево», «плакетки», «розетки» и «линии». Если «кружево», образованное двойным рядом межмембранных частиц, всегда присутствует в основании ресничек инфузорий, то остальные элементы и их сочетания являются характерными признаками различных групп Ciliophora (Bardele, 1981).
В кинетосоме периферические дублеты становятся триплетами: А, Б и С-микротрубочки. Два деривата кинетосом также состоят из микротрубочек: постцилиарная лента микротрубочек обычно связана с задним триплетом 9 кинетосомы, а трансверсальная микротубулярная лента отходит от левого переднего края кинетосомы у триплетов 3 и 4. Третий дериват — поперечно исчерченная фибрилла (кинетодесма), отходит от триплетов 5, 6 и 7. Как правило, кинетиды собраны в продольные ряды — ки- неты, справа от которых располагаются кинетоде- смальные филаменты, направленные к переднему концу тела. Это положение кинетодесмальных дериватов кинетосом было выявлено Шаттоном и Львовым и названо правилом десмодексии (Chatton, Lwoff, 1935). У представителей Karyorelictea и Spirotrichea наибольшее развитие получают постцилиадесмы, располагающиеся также справа от кинет, но направленные к заднему концу тела простейшего. У ряда других таксонов инфузорий преимущественное развитие получают трансверсальные микротрубочки, вытянутые поперек ресничных рядов и начинающиеся с левой стороны кинет. Парасомальные саки обычно располагаются впереди или справа от кинетосом в кинете. Ниже слоя кинетосом у ряда инфузорий могут располагаться митохондрии, иногда образующие характерные скопления вдоль кинет, часть этих органелл располагается в эндоплазме.
Значительное число исследований показало, что устройство соматических кинетид у инфузорий может существенно варьировать. Эти вариации, включающие изменения в размерах, расположении и форме кинето- сомальных дериватов, а также вариации числа кинетосом хорошо совпадают с делением Ciliophora на группы ранга класса (Lynn, 1981; Small, Lynn, 1985; Lynn, Small, 2000; Lynn, 2003). Кинетиды могут быть классифицированы как монокинетиды — одна кинетосома с фибриллярными дериватами; дикинетиды — две кинетосомы с фибриллярными дериватами, и поликинети- ды — более чем две кинетосомы. Последние могут образовывать ряды или скопления, причем фибриллярные дериваты в этих случаях присутствуют не у каждой кинетосомы. Так, в дикинетиде часто лишь одна из кинетосом имеет типичный набор дериватов. Кроме того, представители некоторых групп имеют в кортексе дополнительные фибриллярные компоненты: гименостоматиды — базальные микротрубочки, расположенные вдоль левой стороны кинетосом; филло- фарингиды — субкинетальные микротрубочки, отходящие от основания кинетосом; некоторые гапториды и кольподиды обладают нематодесмами или микроту- булярными тяжами, которые отходят от поверхности пелликулы в глубь цитоплазмы клетки.
Функция самих ресничек представляется очевидной — это органеллы локомоции, способные к сложному (трехплоскостному) движению. Фибриллярные дериваты кинетосомы, по-видимому, играют роль якоря для локомоторной единицы. При этом в случае сильного развития они могут выполнять и определенную роль формообразующих элементов кортекса. Па- росомальный сак, возможно, имеет значение при эндо- цитозе (Nilsson, Deurs, 1983), хотя не всегда ясно, какие вещества могут в норме поглощаться из окружающей инфузорию среды (Lynn, Corliss, 1991).
Очевидно, что кинетом должен выполнять и некоторые сенсорные функции. Долгие поиски в начале XX в. «нейромоториума» у инфузорий или каких-либо специальных субклеточных структур, координирующих их «ощущения», давно забыты. Тем не менее наличие у многих инфузорий укороченных ресничек, не участвующих в локомоции (например, «сенсорные щетинки» гипотрих), с еще пока недостаточно изученной ролью (Gortz, 1982; Lynn, Corliss, 1991) говорит о том, что, возможно, некоторые специализированные реснички у разных инфузорий действительно имеют сенсорные функции. На это указывает и специальный, наиболее богатый состав межмембранных частиц в оболочке специализированных ресничек у гимностоматид, офрисколицид, гетеротрих и олиго- трих (Bardele, 1981).
Общее число кинетид, достигающее, например, у Р. caudatum 15 тыс., сохраняется от деления до деления. Их репликация происходит перед цитокинезом, но для многих инфузорий этот процесс не изучен в деталях. В случае дикинетид родительские кинетосомы разделяются и передняя становится задней во вновь образующемся переднем дублете, в то время как родительская задняя остается в этом же положении в возникающей (дочерней) задней дикинетиде. Некоторые фибриллярные компоненты при этом теряются и возникают de novo у дочерних дублетов. Умножение соматических поликинетид идет более сложно и изучено до сих пор только для Paraurostyla (Jerka-Dziadosz, 1980).
Видоизменения кортекса. Часть соматического кортекса некоторых инфузорий может видоизменяться и служить для прикрепления клетки к субстрату. Наиболее характерно развитие этих приспособлений для симбиотических форм. Как правило, такие видоизменения участка поверхности тела инфузории возникают на переднем (тигмотактильная зона цилиатуры) или заднем (огранеллы прикрепления) концах клетки. Для тигмотакгильных участков кортекса (например, у Соп- chophthirus) изменений компонентов ресничек не отмечено, однако они значительно короче и их инфраци- лиатура устроена иначе (Antipa, 1971). Прикрепление симбиотических форм (Suctoria, Chonotricha, Peritri- cha) часто происходит на стадии бродяжки при помощи специального образования (скопулойд, скопула), которое позднее соединяются с образующимся стебельком (Willey, Walkosz, 1975; Wibel et al., 1997). У свободно- живущих форм прикрепление к субстрату осуществляется различным образом (Hausmann, Hiilsmann, 1996; Hausmann et al., 2003): посредством выделения слизистого материала (Stentor), внутри- и внеклеточными стебельками {Carchesium, Vorticella), пальцевидными выростами {Epistylis и Erastophrya). Прикрепительные аппараты подвижных перитрих и ряда эндобионтных инфузорий работают по принципу присоски, что позволяет не только быстро входить в контакт с хозяином, но и покидать его. Наиболее сложно устроенный аппарат для прикрепления — прикрепительный диск имеют представители отряда Mobilida (Peritrichia) — Trichodina (Hausmann, Hausmann, 1981; Estes et al., 1997).
Экструсомы. Часть кортекса инфузорий, иногда только оральная, иногда лишь соматическая, несет различное число экструсом — пузырьковидных или веретеновидных структур с содержимым, которое может выделяться наружу путем эндоцитоза. В настоящее время для простейших известно около 10 типов этих образований, часть которых встречается и у инфузорий (Rosati, Modeo, 2003). Наиболее хорошо изучены экструсомы некоторых представителей Peniculia — трихоцисты. Этот тип экструсом (размером 2—4 X 0.4—1 мкм), занимающих постоянное место в пелликуле, где их апикальные концы имеют прямой контакт с плазматической мембраной (посредством межмембранных частиц), имеет, по-ви- димому, защитный характер, что экспериментально показано для Paramecium (Harumoto, Miyake, 1991; Knoll et al., 1991; Sugibayashi, Harumoto, 2000).
Содержимое трихоцисты парамеций в покоящемся состоянии представлено паракристаллическим веществом, в состав которого входят (по преимуществу) два белка (Peterson et al., 1987). Форма трихоцист может несколько различаться у разных инфузорий {Paramecium, Frontonia, Disematostoma, Pseudomicrothorax): от бутылко- или амфорообразной до кеглевидной или веретеновидной. Вероятно, что композиция паракристаллического содержимого трихоцист также не одинакова у разных инфузорий. Для трихоцист и мукоцист, характерных для инфузорий из подклассов Peniculia и Hymenosto- matia, показан генезис их из Гольджи-подобных структур в эндоплазме. Сформировавшиеся трихоцисты попадают в кортикальный слой, где они встраиваются в пелликулу. Их число у парамеций примерно втрое меньше, чем количество ресничек. Так, у Р. multimic- ronucleatum около 8000, у Р. caudatum около 5000, а у Р. tetraurelia — около 1000 трихоцист (Hausmann, Allen, 1976; Hausmann, Hiilsmann, 1996; Plattner, 2002).
У парамеций экструзия трихоцист может быть вызвана изменением концентрации ионов в окружающей среде (Plattner et al., 1984). Вершина трихоцисты соприкасается с плазматической мембраной между кортикальными альвеолами посредством кольцевой структуры (розетки слияния), образованной 8—11 межмембранными частицами (Plattner, 2002). Экструзия происходит в два этапа: сначала сливаются плазматическая мембрана и мембрана трихоцисты и затем в течение миллисекунды содержимое экструсомы выстреливает наружу, удлиняясь примерно в десять раз. Сразу вслед за этим мембрана трихоцистной вакуоли отделяется от плазматической мембраны и распадается на мелкие везикулы, которые мигрируют в эндоплазму и используются для синтеза новых экструсом.
Полная замена трихоцист клеткой происходит за время менее 10 ч, что указывает на значимость этих структур. Однако мутанты, лишенные трихоцист, полностью жизнеспособны. Опыты тем не менее показывают, что такие мутанты поедаются хищниками {Di- leptus, Monodinium, Climacostomum) значительно быстрее, чем «нормальные» парамеции (Harumoto, 1994; Miyake, Harumoto, 1996; Sugibayashi, Harumoto, 2000).
Мукоцисты во многом напоминают трихоцисты. Они также имеют паракристаллическое содержимое, состоящее из нитчатых составных элементов. При экструзии это содержимое, однако, разворачивается в трехмерную сеть филоментов, что требует нескольких секунд. Их функции неясны, хотя, предположительно, мукоцисты участвуют в процессе инцистирования, характерного для многих инфузорий и при построении лорики и других защитных «футляров», выделяемых некоторыми Ciliophora.
Токсицисты несомненно служат орудием нападения и защиты. Этот тип экструсом наиболее распространен у хищных инфузорий. При экструзии из капсулы токсицисты выстреливает трубка, либо выварачивающаяся как палец перчатки, либо удлиняющаяся телескопически. Вонзившись в тело жертвы, трубки токсицист механически удерживают добычу и парализуют ее. Соответственно назначению токсицисты располагаются обычно около ротового аппарата (представители классов Litosomatea и Prostomatea). Сходные задачи выполняют гаптоцисты у Suctoria и, возможно, рабдоцисты у некоторых Karyorelictea (Lynn, Corliss, 1991; Rosati, Modeo, 2003).
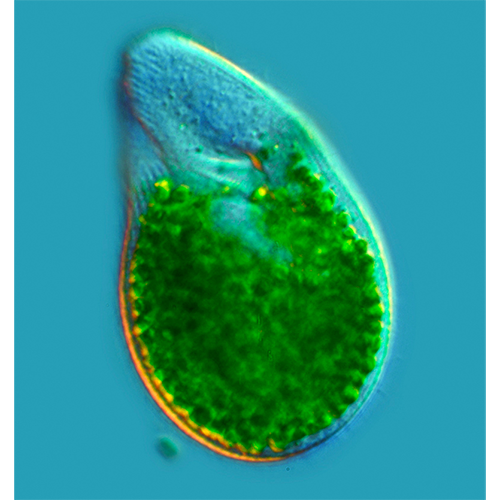
Пигментные гранулы. Некоторые инфузории обладают постоянной окраской, обусловленной присутствием в кортексе специальных пигментоцист или пигментных гранул, локализованных вдоль кинет (рис. 229, а, вкл.; 240, а, вкл.). Пигменты стенторин у Stentor и блефаризмин у Blepharisma, содержащиеся в таких окруженных мембраной гранулах, обеспечивают фоточувствительность этих инфузорий и имеют определенную защитную функцию (Miyake et al., 1990; Miyake, Harumoto, 1995; Miyake et al., 2001).
В состав соматической части кортекса входят также постоянные образования, связанные с комплексом сократительных вакуолей (поры, которые открываются на поверхности кортекса) и с процессом питания инфузорий (цитопрокт, через который происходит дефекация вакуолей с непереваренными пищевыми остатками). В зависимости от видовой принадлежности расположение этих кортикальных структур может быть различным (так же как и детали их устройства), но чаще поры сократительных вакуолей открываются в определенных местах на дорсальной стороне тела, а цитопрокт располагается в задней части инфузории на вентральной ее стороне. Так, у Paramecium и Tetrahymena эти структуры хорошо различимы как на живых объектах (при использовании дифференциального интерференционного контраста), так и при импрегнации кортекса серебром. У парамеций поры сократительных вакуолей открываются в передней и задней трети дорсальной поверхности соматического кортекса (Фокин, 1986, 2002), у тетрахимен — латерально в правой задней части клетки (Nanney, 1980), у F. leucas — в середине дорсальной стороны тела (Foissner et al., 1994).
Поры сократительных вакуолей. Поры сократительных вакуолей могут быть единичными: одна вакуоль — одна пора или множественными — до 3—5 пор на одну вакуоль и соответственно они располагаются в кортек- се (в составе кинет или в межкинетном пространстве) над тем участком эндоплазмы, где локализованы остальные компоненты комплекса сократительной вакуоли (Patterson, 1980; Фокин, 1986; Fokin et al., 1999). Исключение составляют некоторые виды (Ps. dubius, Р. putrinum, Lembadion bullinum), у которых пора приобретает вид извитого выводного протока значительной длины (до 5—10 мкм). У большинства же видов инфузорий поры сократительных вакуолей представляют собой короткий (иногда чуть расширенный к концу) цилиндрический канал, стенки которого армированы спирально скрученными микротрубочками. Основание поры закрывает плазматическая мембранная диафрагма, отделяющая выводной канал от резервуара сократительной вакуоли. Это мембранное образование исчезает при систоле сократительной вакуоли и после выбрасывания порции жидкости, «схлорываясь» восстанавливает изолированность содержимого вакуоли от внешней среды.
Цитопрокт. Цитопрокт — гребневидный удлиненный выступ на пелликуле, который вытянут вдоль тела, начинаясь (у Paramecium) за ротовой зоной кортекса. Около этой кортикальной органеллы альвеолы, эпиплазма и филаментозная сеть прерываются, и эндоплазма соприкасается с плазматической мембраной. У представителей подкласса Choreotrichia цитопрокт отсутствует (Lynn, Small,2000). Наиболее полно эта органелла исследована у Paramecium (Allen, Wolf, 1974). Процесс дефекации занимает лишь несколько секунд. Фагосомы, оказавшиеся в зоне цитопрокта, попадают к нему, видимо, за счет «направляющих» микротрубочек, отходящих от кортекса в глубь эндоплазмы. После слияния мембран пищеварительной вакуоли и плазмалеммы содержимое фагосомы попадает в окружающую клетку среду, а фагосомная мембрана везикулируется на множество пузырьков, возвращаемых в мембранную транспортную систему инфузории.
Эктобионты. С поверхностью соматического кортекса инфузорий нередко связаны эктобионты, в большинстве случаев бактерии. Они наиболее типичны для инфузорий из анаэробных местообитаний (Fenchel, Finlay, 1991). Вероятно, для многих видов, обитающих в местах с дефицитом кислорода, характер такого симбиоза должен быть облигатным, однако специально этот вопрос в большинстве случаев не исследовался. Известен по крайней мере один достоверный случай реального облигатного симбиоза между инфузориями и их эктобионтами — без- ротая кареореликтида Kentrophoros, которая питается за счет бактерий, постоянно присутствующих на дорсальной поверхности ее тела (Fenchel, Finlay, 1989).
Оральный кортекс. Часть кортекса, связанная с ротовыми структурами инфузорий, носит название оральной. Она может быть сильно специализирована в связи с процессом питания, и ее морфологическое строение значительно варьирует в пределах типа Ciliophora. Представители подкласса Suctoria не имеют в дефинитивном состоянии ни ресничек, ни ротового аппарата, а на расселительной стадии бродяжек-томитов обладают только соматической цилиатурой (Довгаль, 1996,2002). Роль рта у этих инфузорий выполняют щупальца, тогда как у большинства остальных инфузорий (кроме Astomatia) всегда имеются оформленные цито- фарингиальные структуры, служащие для захвата пищи. Окружающая рот цилиатура часто тоже оказывается измененной в связи с функцией питания.
Наиболее просто устроены оральные зоны у представителей Litostomatea и Prostomatea, у которых передние кинетиды каждой соматической кинеты дифференцированы в дикинетиды. У литостомей трансверсальные микротубулярные ленты формируют и поддерживают форму цитофарингса (Foissner, Foissner, 1985). У простомат эту роль выполняют постцилиарные микротрубочки (Huttenlauch, Bardele, 1987). Реснички этой зоны могут быть несколько длиннее, чем соматические, а иногда они образуют мелкие по- ликинеты (щетки). Часто у этих инфузорий в состав оральной зоны кортекса входят специализированные экструсомы (Lynn, Corliss, 1991).
Представители класса Nassophorea также имеют оральные дикинетиды и оформленный постцилиарными лентами микротрубочек цитофарингс. Эти инфузории, однако, обладают наиболее сложно устроенной ротовой структурой, называемой корзинкой, или цир- тосом. Микротубулярные тяжи в совокупности с нематодесмами облегчают прохождение пищи в цитоплазму (Hausmann, Peck, 1979). Предротовые структуры у представителей этой группы оформлены как поликинетиды. Когда оральная зона образует углубление (глотку), как например у представителей Peniculia — Paramecium, Frontonia или Aeobursaridium, поликинеты, именуемые в данном случае пеникулюсами и квадрилюсом, оказываются погруженными. Биение их ресничек создает ток, увлекающий в глотку к ротовому отверстию бактерий или других инфузорий, служащих пищей.
Олигохименофоры являются преимущественно бактериофагами-фильтраторами. Система олигохиме- ниума: 3 продольные поликинеты на левой стенке глотки и пароральная (эндоральная) мембрана, расположенная под углом к ним на правой, создает оптимальные условия для захвата и фильтрации из окружающей среды бактерий и других мелких объектов. Реснички поликинет осуществляют направленный ток воды, а пароральная мембрана служит фильтром, задерживающим бактерий в глотке. В зоне ротовых поликинет отсутствуют альвеолы, а расположение пара- сомальных саков и филаментозных дериватов кинетосом отлично от соматической зоны кортекса (Lynn, Corliss, 1991).
Спирально закрученная адоральная зона поликине- тид у Spirotrichea настолько заметна, что она дала название всему классу. Эти инфузории, используя оральные реснички, одновременно создают ток воды к ротовому отверстию и фильтруют находящиеся в воде мелкие пищевые объекты (Fenchel, 1986).
Долгое время крупные кольподиды Bursaria и Woodrujfia, также имеющие адоральную зону полики- нетид, причислялись к спиротрихам (Corliss, 1979). На самом деле структура соматических кинетид (в том числе и переходной зоны ресничек) различна в этих двух классах Ciliophora, и на этом основании упомянутые виды были перемещены в Colpodea (Lynn, Corliss, 1991; Foissner, 1993; Фокин, 1995). Кроме того, характер фильтрации воды в ротовом аппарате кольподид больше напоминает Oligohymenophora, чем Spirotrichea (Fenchel, 1986).
Среди представителей Phyllopharyngea имеются виды с сильно различающимся устройством ротовой зоны. У хонотрих, очевидно, произошло простое перемещение части соматической цилиатуры, с изменением ее позиции на 180°. В отрядах Chlamydodontida, Disteriida и Hypocomatida большинство инфузорий обладают корзинковидным цитофарингсом, или цирто- сом, весьма напоминающим ротовой аппарат Nassophorea (Corliss, 1979). Наконец, как было уже отмечено, Suctoria не имеют рта, и питание осуществляется у этих инфузорий через специализированные пищевые щупальца, обладающие сложным микротубулярным «скелетом».
Безротые инфузории, относящиеся в подавляющем большинстве к подклассу Astomatia класса Oligohy- menophorea, по-видимому, питаются эндоцитозом, с использованием парасомальных саков, или пиноци- тозом, через пелликулу. Вопрос этот до сих пор не изучен, но наблюдения над безротыми мутантами Tetra- hymena (Rasmussen, Orias, 1975) говорят в пользу существования таких возможностей. Особый случай питания безротой инфузории за счет эндоцитоза экто- бионтных бактерий, постоянно присутствующих на дорсальной стороне клетки инфузории, описан для представителя Karyorelictea — Kentrophoros (Centro- phorelld) (Faure-Fremiet, 1950a; Fenchel, Finlay, 1989).
Домики. Помимо кортекса, имеющего в том числе и защитные функции, некоторые инфузории выделяют на поверхность тела постоянные защитные образования типа лорики или футляра. Чисто гиалиновая или армированная (агглютинированная) природными частичками лорика присутствует у всех представителей отряда Tintinnida (Spirotrichea) и у некоторых Hetero- trichea: Folliculina, Stentor. Виды, имеющие лорику или футляр, встречаются и в других группах инфузорий, но, по-видимому, не среди литостомей и нассофо- рей (Lynn, Corliss, 1991). Развитие лорики изучено пока лишь для нескольких форм, в том числе для Eufol- liculina (Mulisch, Hausmann, 1983). Для этой инфузории было показано, что в секреции лорики участвуют мукоцистоподобные тельца, расположенные в кортексе инфузорий.
ЭНДОПЛАЗМА И ЕЕ ВКЛЮЧЕНИЯ
Основные органеллы и системы клетки инфузорий расположены в эндоплазме. Наиболее заметные из них, особенно при светооптическом микроскопировании, — элементы ядерного аппарата, пищеварительные и сократительные вакуоли. Как уже было отмечено, присутствие в клетке инфузорий ядер двух типов — соматического (макронуклеус) и генеративного (микронуклеус) — считается неотъемлемой особенностью типа Ciliophora. До сих пор не найдено одноклеточных, принадлежащих к этому типу, у которых отсутствовал бы ядерный гетероморфизм. Некоторое время таковым считался Stephanopogon, имеющий несколько одинаковых ядер (Raikov, 1969; Corliss, 1979), но, по современным данным, это простейшее принадлежит к гетеротрофным жгутиконосцам (Lipscomb, Corliss, 1982; Patterson, Brugerolle, 1988). Макронуклеусы, как правило, очень крупные ядра, богатые ДНК, делящиеся амитотически. У некоторых инфузорий соматическое ядро представлено в вегетативных клетках множеством фрагментов, а у кариореликтид макронуклеусы сравнительно мелкие, содержат мало ДНК (парадип- лоидные) и не способны делиться. Потеря макронуклеуса для инфузории всегда детальна, тогда как клетки, лишенные микронуклеуса, могут при определенных условиях существовать и даже давать жизнеспособные амикронуклеарные линии (Fokin, Ossipov, 1981; Ng, 1986).Микронуклеус. Микронуклеусы — мелкие, обычно шаровидные ядра, редко превышающие 10 мкм в диаметре. Если инфузория имеет 1 или 2 микронуклеуса, то часто они располагаются рядом с соматическим ядром или даже в его углублении, хотя у многих (особенно крупных) инфузорий, которые имеют множественные (до 10 и более) генеративных ядер, они могут находиться в различных местах эндоплазмы. У Karyorelictidae ядра часто образуют комплексы (группы) сближенных ядер, в состав каждой из которых входят 2 или несколько макронуклеусов и 1 микронуклеус (Райков, 1978). В редких случаях — у представителей Cyrtolophosidida (Colpo- dea) — микронуклеус оказывается включенным в перинуклеарное пространство макронуклеуса и ядер- ный комплекс, имеющий общую оболочку, становится неразрывным (Foissner, 1993; Raikov, 1996). Обычно же микронуклеус инфузорий имеет типичную ядер- ную оболочку, несущую значительное число пор.
Хроматин микронуклеусов в большинстве случаев сильно конденсирован й представляет собой одно или несколько плотных хроматиновых тел, занимающих большую часть объема ядра (рис. 246, вкл.; 247, вкл.). Тем не менее морфология этих ядер у разных инфузорий, часто даже близкородственных, может существенно различаться. Например, среди видов рода Paramecium есть обладающие генеративными ядрами «компактного», «хромосомного», «эндосомного» и «везикулярного» типа (Fokin, 1997). У некоторых представителей Кагу- orelictae имеются «спермальные» микронуклеусы (Raikov, 1996).
Кариоплазма генеративных ядер помимо хроматина может содержать белковые тела, кристаллоиды, центры организации микротрубочек, микрофиламен- тозные агрегаты (Raikov, 1996; рис. 248, вкл.). Изредка кариоплазму и перинуклеарное пространство микронуклеуса могут заселять бактериальные эндобионты.
Деление микронуклеусов инфузорий происходит по типу внутриядерного (закрытого) ортомитоза (Райков, 1978). Перед делением хромосомы ядра конденсируются и образуют метафазную пластинку. Аппарат веретена деления в это время становится заметным, а форма микронуклеуса, изменяясь, становится эллипсоидной (рис. 248, а, в, вкл.). В веретене деления генеративного ядра выявляются микротрубочки двух родов: непрерывные (тянущиеся от одного полюса ядра до другого) и связанные с кинетохорами хромосом (заканчивающиеся на одном из полюсов). Позиция кине- тохоров и детали их морфологии могут несколько различаться у разных инфузорий. Иногда они не одинаковы даже в пределах родовой группы (Paramecium} (Lewis, 1975). На стадии анафазы—телофазы происходит наибольшее удлинение ядра, приводящее к тому, что дочерние микронуклеусы оказываются на противоположных концах делящейся клетки. Это необычно длинное веретено деления обеспечивает гарантированное попадание генеративного ядра в каждую из дочерних клеток, а при реорганизации ядерного аппарата в ходе конъюгации — попадание зачатков макро- и миронуклеусов в концевые участки цитоплазмы, что необходимо для их последующей диференцировки (Yang, Takahashi, 2002).
Ядерная оболочка, интактная на всех стадиях кариокинеза, может непосредственно становиться оболочкой дочерних микронуклеусов (Paramecium, Nassula, Didinium и многие другие виды). У ряда видов: Dilep- tus, Blepharisma, Stentor — под внутренней мембраной делящегося ядра образуется новая оболочка дочерних ядер, после чего двойная мембрана «родительского» ядра распадается (Райков, 1978; Осипов, 1981).
Как генеративное ядро микронуклеус способен к двуступенчатому мейозу. Характерной особенностью профазы I мейотического деления микронуклеуса многих инфузорий является своеобразная стадия «серпа», когда линейные размеры ядра значительно увеличиваются и оно приобретает соответствующую форму. У некоторых инфузорий эта стадия отсутствует, а про- фазное ядро напоминает «парашют». Нередко инфузории, принадлежащие к одной группе (Paramecium, Frontonia), имеют разную морфологию профазного микронуклеуса (Fokin et al., 2001).
Хромосомы микронуклеуса обычно мелкие и многочисленные — гаплоидный набор от 16 до 150, хотя у Tetrahymena генеративное ядро имеет лишь 5 пар хромосом. У некоторых видов известна вариация числа хромосом между клонами. Для Р. bursaria, Р. putrinum и Р. caudatum показано, что генеративное ядро не является диплоидным, каким оно обычно считается у инфузорий, а представляет собой низкополиплоидное образование (Chen, 1940; Jankowski, 1972; Осипов, 1981; Fokin, 1997).
В составе кариоплазмы микронуклеусов не выявляются ядрышкоподобные структуры (рис. 247, 248, вкл.) и транскрипционная активность этого ядра принимается близкой к нулю. Соответственно этот тип ядер инфузорий обычно считался инертным в вегетативной жизни клетки. Однако за последнюю четверть века накопилось много данных о роли генеративного ядра в процессах морфогенеза и особенно стоматоге- неза различных инфузорий (Фокин, 1978, 2002). Экспериментальное удаление микронуклеуса у различных инфузорий (Paramecium, Tetrahymena, Euplotes, Pseu- dourostyla, Stylonychia) всегда приводит к нарушениям нормального хода стоматогенеза и функционирования ротового аппарата инфузорий, а также общего морфогенеза клетки (Фокин, 1983а, 19836; Ng, 1986; Tam, Ng, 1986; Takahashi, 1988; Lu, Ng, 1991; Oka, Mikami, 1996; Фокин, 2002). При этом даже в пределах рода Paramecium существует некоторая видовая специфика этих функций микронуклеуса и связанных с ними клеточных процессов (Фокин, 1983а, 19836; Tam, Ng, 1986).
Стоматогенетическая роль генеративного ядра в ходе ядерной реорганизации при конъюгации инфузорий хорошо известна (Осипов, Тавровская, 1969; Ng, 1986; Chau, Ng, 1989). Таким образом, очевидно, что некоторые (единичные) гены микронуклеуса активны на протяжении всего жизненного цикла инфузорий. В этом смысле противопоставление морфогенетической роли генеративного ядра в половой и вегетативной части жизни клетки, которое обычно подчеркивается в литературе, оказывается неоправданным. Эта роль должна быть распространена на весь жизненный цикл инфузорий.
Макронуклеус. Соматическое ядро инфузорий — макронуклеус, в вегетативной жизни транскрипцион- но активно, обычно называется полиплоидным. На самом деле организация этого типа ядер у инфузорий значительно варьирует в разных группах и, как правило, амплификации подвергается не весь геном ядра, а лишь его часть (иногда весьма незначительная, как у гипотрих). Еще большее разнообразие имеют морфология макронуклеуса, его размеры и форма. В зависимости от видовой принадлежности макронуклеус инфузорий может быть сферическим, удлиненным, колбасовидным, лентовидным, бобовидным, кустистым, четкообразным или разделенным на две и более части (Райков, 1978; Raikov, 1996). Размеры соматического ядра, по-видимому, находятся в соответствии с величиной клетки инфузории, и у крупных форм (Bursaria, Spirostomum) макронуклеус может достигать длины 2 - -3 мм.
Ядро окружено типичной ядерной оболочкой с многочисленными порами (рис. 251, вкл.), число которых может существенно варьировать, но обычно (Paramecium, Tetrahymena) достигает 100 и более на 1 мкм2. Внешняя мембрана оболочки макронуклеуса часто контактирует с эндоплазматическим ретикулумом и иногда с оболочкой микронуклеусов (Raikov, 1969; Осипов, 1981; Raikov, 1996).
При светооптическом наблюдении макронуклеус кажется заполненным достаточно плотным хроматином, как правило, дающим интенсивную окраску на ДНК, выявляемую по методу Фельгена. Электронномикроскопические исследования этого ядра инфузорий показывают, что в его матриксе расположено обычно большое число мелких хроматиновых тел (0.1—0.5 мкм диаметром), часто аностоматизирующих между собой. Помимо хроматина макронуклеус всегда содержит значительное число нуклеол (часто весьма крупных — до 1—2 мкм), состоящих из гранулярного и фибриллярного компонентов различной конфигурации и площади. У некоторых видов постоянно (или в определенном физиологическом состоянии) большинство нуклеол расположено вплотную к внутренней мембране ядерной оболочки. Размеры, структура и положение нуклеол как структур очень динамичных зависят от состояния клетки и условий внешней среды.
В качестве дополнительных элементов (включений) в макронуклеусе могут встречаться плотные или рыхлые фиброзные белковые сферы (Blepharisma, Hal- teria, Lacrimaria, Stentor), актиновые микрофиламенты и микротрубочки (Stylonychia, Tetrahymena, Paramecium, Acineta) и нередко бактериальные эндобионты. Значительно реже — в макронуклеусе некоторых инфузорий — Р. putrinum, Eupletes sp. и Paraholos- ticha sterkii могут поселяться жгутиконосцы (Gortz, 1996).
У некоторых инфузорий (Phyllopharyngea) макронуклеус дифференцирован на две части (два кариоме- ра): ДНК-богатый ортомер и ДНК-бедный парамер (Янковский, 1973; Radzikowski, 1985). Ортомер может окружать парамер, как это имеет место у Chilodonella. Если ортомер содержит многочисленные плотные хроматиновые тела и нуклеолы, то парамер состоит лишь из эндосомы (конденсированного ДНК-содержащего тела). Функциональное значение гетеромерии остается пока неясным. Репликация ДНК проходит в макронуклеусе в течение интерфазы и занимает значительное время (Raikov, 1996). Для большинства инфузорий этот процесс идет асинхронно, однако у Hypotrichida и части Oligo- Irichida репликация ДНК происходит локально (полоски репликации), волнообразно перемещаясь вдоль ядра. Как правило, репликации предшествует компактизация ядра или слияние его частей (в случае существования нескольких фрагментов).
Деление макронуклеуса инфузорий (за исключением соматических ядер Karyorelictea, которые вообще не делятся) происходит амитотически (Raikov, 1978, 1996). Оно не включает конденсацию хромосом и не связано с качественным изменением и точной сегрегацией содержимого ядра. Характерно, что микротрубочки, обычно появляющиеся в кариоплазме на стадии удлинения макронуклеуса, не связаны с хроматином и служат, по-видимому, именно для вытягивания ядра. У представителей Heterotrichida микротрубочки, также сопровождающие деление макронуклеуса, располагаются вне ядра (Jenkins, 1977). У некоторых гипотрих (Stylonychia) перед началом вытягивания макронуклеуса происходит интенсивная ротация кариоплазмы, механизм которой пока не известен. Функционально такое «перемешивание» содержимого ядра способствует, по-видимому, эквивалентному распределению хромосомного материала между дочерними макронуклеусами. После вытягивания делящийся макронуклеус образует два, как правило равных, компактных продукта (дочерних ядра). Далее они приобретают форму и размеры нативного макронуклеуса (удлиненный, четковидный, фрагментированный и т. д.). В ряде случаев (Suctoria, Astomatida) возможно также неравное деление или почкование макронуклеуса, причем оно может быть единичным или множественным. Быстрое развитие нового соматического ядра из почки или мелкого фрагмента (одного сегмента четковидного ядра у Stentor или Spirostomum) представляет собой регенерацию и показывает, что даже незначительная часть макронуклеуса эквипотентна целому ядру (Фокин, Сково- родкин, 1991).
Несмотря на элиминацию части генома микронуклеуса (при развитии макронуклеуса), в некоторых случаях возможно говорить о распределении геномов в ходе деления соматического ядра (Colpodidae, Нуше- nostomatia). Так, у Tetrahymena делящийся макронуклеус содержит около 90 хроматиновых кластеров, соответствующих, по-видимому, геномным наборам (Raikov, 1995; Nilsson, 2000). Показано, например, что содержание ДНК в таких агрегатах у Tillina соответствует диплоидному хромосомному набору (Frenkel, 1982).
Количество ДНК в макронуклеусах инфузорий (на стадии GJ варьирует от 1.4 пг (Uronema) до 38 000 пг (Bursaria). Соматическое ядро Т termophila, например, содержит 10 пг ДНК, разные Paramecium — от 70 до 300, крупные виды Stylonychia — около 1000, и Stentor — 3000 пг (Raikov, 1996). Таким образом, количество ДНК макронуклеуса в целом оказывается пропорционально размерам инфузории. Количество ДНК в макронуклеусах во много раз превышает этот показатель для дипйоидного генеративного ядра: в 23 раза у Т. termophila, в 430 раз у инфузорий группы Р. aurelia и в 6500 раз у Spirostomum (Райков, 1978), что и служило поводом говорить о полиплоидности соматического ядра инфузорий (Райков, 1967).
Сравнение нуклеотидных последовательностей ДНК макронуклеуса и микронуклеуса показало уменьшение (для ряда видов значительное) размера генома нативного соматического ядра. Это означает, что при развитии макронуклеуса происходит редукция ряда последовательностей (Steinbriick, 1983, 1986; Ammer- mann, 1985). Если для Tetrahymena такие потери составляют 10---15 % (Gorovski, 1980; Karrer, 2000), то у Stylonychia при развитии нового макронуклеуса теряется свыше 95 % генома микронуклеуса (Steinbriick, 1990). Таким образом, нативный макронуклеус у многих видов инфузорий состоит по большей части из многочисленных копий лишь некоторых генов или их комплексов. Поскольку амплификация части генетического материала не соответствует классическому понятию полиплоидии, Шварцем (Schwartz, 1978) для макронуклеусов инфузорий был предложен термин «амплиплоидия», что в большей степени соответствует современным данным о генетической природе соматического ядра Ciliophora (Raikov, 1996).
Данные биохимии, электронной микроскопии и особенно результаты пульсэлектрофореза показывают, что макронуклеус инфузорий не содержит целых хромосом. Очевидно, при развитии макронуклеуса хромосомы фрагментируются, какие-то их части утрачиваются или перестраиваются (Blackbum, Karrer, 1986; Raikov, 1996). По-видимому, возможны два пути реорганизации генома соматического ядра инфузорий. В первом случае макронуклеус содержит достаточно длинные молекулы ДНК, представляющие собой фрагменты хромосом (субхромосомы). Каждый из таких фрагментов содержит несколько или даже много генов. Так дело обстоит у Tetrahymena и Paramecium (Karrer, 1986, 2000; Preer, 1989; Steinbriick, 1990). Возможно, что эта модель реализуется также у Stentor, Blepharisma, Bursaria и Colpoda (Raikov, 1996). Другой путь наиболее типичен для гипотрих: макронуклеус содержит лишь короткие фрагменты ДНК (минихромосомы), соответствующие одному гену (Steinbriick, 1990; Bleyman, 1996; Raikov, 1996). Нечто подобное происходит у некоторых олиготрихид и циртофорид, например у Halteria и Chilodonella (Metenier, Huf- schmid, 1988). Подсчеты показывают, что базовый геном макронуклеуса гипотрих (Oxytricha, Stylonychia и Euplotes) состоит из 10—40 тыс. минихромосом. Принимая во внимание, что количество копий этих образований также весьма значительно, можно говорить о миллионах минихромосом в одном макронуклеусе.
Митохондрии. Инфузории не нуждаются в специальной дыхательной системе, поскольку кислород и углекислый газ свободно диффундируют через пелликулу клетки. Однако большинство этих простейших имеют митохондрии — органеллы дыхания, свойственные всем эукариотам и содержащие типичный набор ферментов: цитохромов и цитохром-оксидаз (Blum, 1996). Хорошо заметные митохондрии (1— 3 мкм) располагаются в эндоплазме, хотя обычно они в большом числе входят и в состав кортекса. Совокупность митохондрий именуется хонд- риомом, который в некоторых случаях имеет строго определенную клеточную топографию. Число митохондрий значительно варьирует у разных инфузорий и, по-видимому, зависит от размеров клетки, видовой принадлежности и экологии этих простейших. Так, у представителей комплекса Т. pyriformis около 1500 митохондрий, у Paramecium — в несколько раз больше.
Матрикс митохондрий отделен от цитоплазмы двойной мембраной. Внутренняя мембрана образует многочисленные выросты (кристы), заполняющие объем органеллы. Характерной чертой митохондрий инфузорий считается присутствие в них только трубчатых крист, которые могут иметь разную упорядоченность и размеры, но никогда не переходят в пластинчатые, дисковидные или пузырьковидные (Lynn, Corliss, 1991; Серавин, 1993; Фокин, 1993а; Фокин и др., 1993; Paulin, 1996; Карпов, 2001). Помимо крист в матриксе митохондрий иногда обнаруживаются кристаллические или фибриллярные включения. Крайне редко структура митохондрий (видимо, в связи с физиологическим состоянием клетки) может быть нарушена (Yusa, 1964), в том числе и в результате инфекции этих органелл бактериоподобными симбионтами.
В анаэробных условиях значительная часть представителей 8 отрядов инфузорий (Armophorida, Entodi- niomorphida, Haptorida, Heterotrichida, Odontostomati- da, Plagiopylida, Thigmotrichida и Vestibuliferida) утратила митохондрии и приобрела их функциональный аналог — гидрогеносомы (Muller, 1988; Fenchel, Finlay, 1991, 1995). Гидрогеносомы—небольшие пузырьки (0.5—1.5 мкм), ограниченные двойной мембраной, содержат ферменты, окисляющие пируваты и малаты, с освобождением молекулярного водорода. Обычно эти структуры рассматриваются как дериваты утраченных в эволюции митохондрий (Карпов, 2001).
Другие мембранные органеллы. Помимо митохондрий или гидрогеносом в эндоплазме инфузорий встречается значительное число других мембранных органелл и структур, характерных для эукариотной клетки (Allen, 1988; Paulin, 1996). Среди них пероксисомы — мелкие тельца округлой формы, ограниченные мембраной и содержащие оксидазы; лизосомы (первичные и вторичные) — мелкие вакуолеобразные тельца (0.5—0.8 мкм), содержащие литические ферменты и играющие исключительную роль в клеточной защитной системе, системе пищеварения и транспорта (Allen, Fok, 1993; Blum, 1996; Radek, Hausmann, 1996). Мембранные компоненты эндоплазмы представлены эндоплазматическим ретикулумом (как гладким, так и содержащим многочисленные рибосомы) и его дериватами: аппаратом Гольджи (диктиосомами) и мембранными составляющими системы сократительных вакуолей.
Включения. В эндоплазме, как правило, содержится некоторое количество продуктов метаболизма клетки в виде капель жира, гликогена и кристаллов (кристаллоидов). В зависимости от физиологического состояния и видовой принадлежности эти включения могут быть более или менее многочисленными. Кристаллы, обнаруженные в цитоплазме инфузорий, всегда заключены в вакуоль и состоят прежде всего из солей кальция и фосфора. Также в виде включений у инфузорий отмечены соли бария (Loxodes) и магния (Lynn, Corliss, 1991; Hausmann, Hiilsmann, 1996; Grover et al., 1997; Creutz et al., 2002). Считается, что конкреции солей кальция в цитоплазме Spirostomum играют функциональную роль в процессах сокращения—вытягивания у этой инфузории (Vivier et al., 1969).
Эндобионты. Весьма обычным явлением можно теперь считать присутствие в эндоплазме (а иногда и в кортексе) инфузорий эндобионтных микроорганизмов. Это могут быть другие инфузории (Suctoria), водоросли, грибы, дрожжи, микроспоридии и наиболее часто бактерии (Heckmann, Gortz, 1991; Фокин, 1993а; Fokin, 2004а; Gortz, 1996, 1998). Так, для представителей рода Paramecium описано свыше 30 различных бактерий, могущих заселять цитоплазму инфузорий (Heckmann, Gortz, 1991; Фокин, 1993а; Fokin, 2004а). Как правило, бактериальные эндобионты не могут жить вне тела хозяина, и в этом смысле они безусловно являются облигатными. Инфузории же, за исключением их части, приуроченной к анаэробным местам обитания, некоторых видов рода Euplotes, Parauronema acutum и, возможно, Climacos- tomum, могут существовать без каких-либо внутриклеточных бактерий (Фокин, 1993 а; Fokin, 2004а). Достаточно часто в природе можно обнаружить «зеленых» инфузорий, в цитоплазме которых постоянно поддерживаются водоросли, наиболее типичны — Chlorella sp. В ряде случаев возможно поддержание в цитоплазме Chiliophora и пластид, попавших в хозяина в составе пищевых объектов (водорослей, жгутиконосцев и т. д.) и стабилизировавшихся в цитоплазме инфузорий в качестве эндобион- тов (Laval-Peuto, Febvre, 1986).
Фагосомы. Значительное число пищеварительных вакуолей (фагосом) и ассоциированных с ними структур обычно наблюдается в клетках физиологически активных инфузорий, обладающих фаготрофным типом питания. По пищевым предпочтениям инфузории делятся на целый ряд групп (см. общую характеристику типа), и соответственно в фагосомах можно обнаружить самые разные включения. Считается, что пиноцитоз возможен у инфузорий через парасомальные саки (Radek, Hausmann, 1996).
Многие Ciliophora — бактериофаги. Образование фагосом и дальнейший пищеварительный цикл наиболее подробно были изучены именно для этих Cilio- pliora (Paramecium, Tetrahymena'). Очевидно, в общих чер тах он сходен у всех инфузорий. Фагосомы образуются в цитостоме, в конце глоточного углублении, за счет замыкания пойманных бактерий (пли других пищевых объектов) в мембрану. Эта мембрана происходит от дискоидальных пузырьков (0.3 мкм диаме тром), во множестве аккумулирующихся на эндоплазматической поверхности так называемой «ребристой стопки» глотки парамеции. Подобные структуры были обнаружены также у Climacostomum, Eu/blliculina и Euplotes (Radek, Hausmann, 1996).
Было рассчитано, что клетка парамеции затрачивает около 16 000 мкм2 мембран для образование фагосом в течение 20 мин питания. Очевидно, что эти мембраны не возникают de novo, а мигрируют по транспортной системе клетки от уже сформировавшихся пищеварительных вакуолей (при уменьшении их объема на II стадии пищеварения) и (или) из зоны цитопрокта. Рециклирующие мембраны разной конфигурации обнаружены у всех исследованных в этом отношении инфузорий (Fok, Allen, 1988; Lynn, Corliss, 1991; Allen, 1995; Radek, Hausmann, 1996).
Процесс пищеварения у инфузорий включает по крайней мере 4 стадии: I — образование фагосом, II — их закисление — конденсацию, III — слияние с лизосомами, с последующим перевариванием содержимого вакуолей, VI — дефекацию. На стадии закисления (сразу после образования пищеварительной вакуоли) фагосомы сливаются с ацидосомами, что приводит к резкому снижению значений pH и созданию оптимальной среды для действия литических ферментов (для Tetrahymena ацидосомы не найдены). Образованные вакуоли за счет циклоза, со скоростью примерно 3 мкм/с, перемещаются по эндоплазме. В процессе движения, вероятно, принимают участие актомиозиновая система и микротрубочки, связанные с цитофарингсом. В течение нескольких минут на поверхности фагосом образуются выросты, отшнуровывающиеся в цитоплазму, что приводит к компактизации фагосомы и трансформации свойств ее мембраны (Allen, Fok, 1983). Далее вокруг фагосомы накапливаются лизосомы и через 7— 8 мин после начала циклоза они синхронно сливаются с ее мембраной, трансформируя вакуоль в фаголизосому . Комплекс энзимов, входящий в состав лизосом, переваривает пищу, а от поверхности вакуоли снова отшнуровывается значительное число пальцевидных выростов — рециклирующих лизосом . Пищеварение завершается за 20—25 мин. После этого фагосомы оказываются способными вследствие очередного изменения их мембраны к взаимодействию с микротрубочками, направляющими их к цитопрокту. Процесс дефекации описан в разделе, посвященном кортексу.
Сократительные вакуоли. Работа системы осморегуляции, поддерживающей определенный баланс жидкости и солей внутри клетки инфузорий (особенно пресноводных), направлена на удаление излишней воды, которая проникает извне, и регуляцию ионного состава цитоплазмы с помощью различных ионных помп и каналов. У большинства инфузорий она представлена в эндоплазме комплексом одной или нескольких (многих) сократительных вакуолей (сферических образований переменного диаметра), включающих (в наиболее полном виде) спонгиом, систему собирающих каналов (канала) и центральную ампулу. Вакуоль сравнительно медленно заполняется жидкостью (диастола) и периодически быстро опорожняется во внешнюю среду (систола), вследствие чего ее размер изменяется во времени в среднем от 2 до 20 мкм (в зависимости от видовой принадлежности). Частота пульсаций сократительных вакуолей у разных инфузорий не одинакова и, по-видимому, зависит от размера клетки простейшего и осмомолярности окружающей среды: у мелких форм сокращения более часты, чем у крупных, а у некоторых морских инфузорий — очень редки. Для части морских форм сократительные вакуоли вообще не обнаружены.
Относительно движения воды в цитоплазме инфузорий нет однозначного мнения. Вероятно, и отток жидкости из эндоплазмы в систему сократительных вакуолей осуществляется комбинацией нескольких пассивных (физических) и активных (биохимических) процессов. Спонгиом инфузорий, осуществляющий первичный сбор жидкости, представляет собой комплекс постоянно присутствующих в цитоплазме «жестких» трубочек шириной около 60 нм и системы мембран гладкого эндоплазматического ретикулума (лакун и пузырьков), связанной с просветом собирающего канала (каналов). Этот комплекс у Paramecium представляется наиболее сложным. У большинства видов рода имеется несколько (от 5 до 16) приводящих, радиально расположенных, каналов, связанных с собственными спонгиомными участками. Именно там происходит выход жидкости в просветы каналов, с обратным транспортом большинста ионов, содержащихся в растворе. Скапливаясь некоторое время в каналах, жидкость из них поступает (обычно синхронно) в центральную ампулу, что приводит к значительному увеличению объема последней (Patterson, 1980). Для ряда инфузорий наличие специальных собирательных каналов не выявлено, иногда такие отличия демонстрируют близкородственые виды (представители родов Paramecium и Frontonia).
Характер сокращения объема центральной ампулы сократительной вакуоли инфузорий дает основание думать, что оно осуществляется активно за счет сократимых элементов, окружающих центральную ампулу. В случае инфузорий рода Paramecium определенную роль в этом процессе могут играть ленты микротрубочек, армирующие приводящие каналы и с них переходящие на стенки центральной ампулы. Далее эти микротубулярные ленты в виде протуберанца сходятся к поре сократительной вакуоли, образуя в ее стенке плотную спираль (Jurand, Selman, 1969). Аналогичные микротубулярные ленты выявлены и для Р. duboscqui, не имеющей в сократительных вакуолях приводящих каналов (Фокин, 1986). Структура и топография пор сократительных вакуолей, входящих в состав кортекса инфузорий, обсуждались ранее в соответствующем разделе.
МОРФОГЕНЕЗ
В ходе жизненного цикла многие инфузории могут значительно менять свой облик, при этом некоторые органеллы или их комплексы исчезают или появляются заново, увеличиваясь или уменьшаясь в числе. Процесс закономерной трансформации морфологии и состояния клетки простейших в целом называют морфогенезом (Hausmann, Hiilsmann, 1996). Одним из наиболее регулярно повторяющихся морфогенетических процессов в ходе жизненного цикла инфузорий считается процесс клеточного деления. Деление клетки инфузорий затрагивает самые разнообразные стороны ее организации, и в сравнительном аспекте его изучение дает много ценной информации о таксономических и филогенетических связях в пределах группы. Помимо кариокинеза, кратко рассмотренного выше, особенное значение имеет кортикальный сомато- и стоматогенез, в ходе которого дочерние клетки восстанавливают характерный набор клеточных компонентов и план строения своего тела (Гераси- мова, 1989; Lynn, Corliss, 1991; Lynn, 2003).Деление. Деление у инфузорий может происходить в активной фазе или в цисте размножения и может быть моно- или политомическим (Foissner, 1996). При монотомическом делении, которым обладает большинство инфузорий, в результате деления образуются две дочерние особи: протер — передняя и опистер — задняя. Палинтомическое деление (палин- томия, стробиляция или почкование) приводит к появлению нескольких (многих) дочерних особей, как правило отличающихся от материнской клетки меньшими размерами. Этот тип деления характерен для апостомат, астомат, хонотрих и сукторий. Монотомическое деление может быть трансверсальным (деление вдоль главной оси тела, что происходит у большинства инфузорий), энан- тиотропическим (при котором происходит некоторое отклонение оси деления от главной оси тела — Oligotrichia) или продольным (когда оси продуктов деления располагаются параллельно — Peritrichia).
Перед завершением клеточного деления в норме все компоненты тела инфузории должны быть удвоены. Для соматических структур это прежде всего кинетосомы с их фибриллярными дериватами, поры сократительных вакуолей и цитопрокт. Образование новых кинетосом происходит обычно в связи с предсу- ществующими, и этот процесс, начавшись еще в интерфазный период, интенсифицируется у болыпинста инфузорий перед стоматогенезом. Новые поры сократительных вакуолей также появляются перед самым процессом деления. У большинства инфузорий, обладающих 1 терминальной сократительной вакуолью (Tetrahymena), ее сохраняет задний продукт деления. Если сократительных вакуолей две, то обе дочерние клетки получают по одной предсуществующей поре и одну достраивают заново (Paramecium). Сходным образом воссоздается в ходе деления цитопрокт: поскольку обычно он располагается в задней части клетки, передний продукт деления (протер) должен образовать эту органеллу заново. В случае сидячих форм (Chonotricha) органоид прикрепления также образуется заново у протера.
Кортикальный стоматогенез в ходе клеточного деления — одна из наиболее интенсивно изучаемых проблем цилиатологии. Как уже было отмечено, особенности стоматогенеза позволяют систематикам выявлять филогенетические связи между группами инфузорий, что и определяет повышенный интерес исследователей к этой проблематике (Corliss, 1979; Puytorac et al., 1994; Eisler, Fleury, 1995). Кроме того, изучение особенностей стоматогенеза инфузорий (и их морфогенеза вообще) составляет предмет исследования биологии развития как удобный пример субклеточной дифференцировки (Nanney, 1980; Frankel, 1989). В качестве примера использования данных по стоматоге- незу можно привести историю помещения хонотрих в класс Phyllopharyngea вместе с циртофоридами. Основываясь на морфостатических характеристиках, было бы трудно распознать связь этих значительно различающихся групп (Lynn, Corliss, 1991). Однако исследование развития ротовых и соматических частей кортекса при почковании хонотрих ясно показало их значительное сходство с циртофоридами (Guilcher, 1951; Янковский, 1973).
Стоматогенез. Представители каждого класса инфузорий в целом обладают идентичным типом стоматогенеза. Последовательность событий при развитии нового ротового аппарата инфузорий в значительной мере сходна. Сначала появляется зачаток структуры, располагающийся обычно в районе экватора материнской клетки. Далее начинается пролиферация кинетосом, входящих в ротовые структуры. Она происходит в большинстве случаев в связи с предсуществующими кинетами. У гипотрих, некоторых олиготрих и энтоди- ниоморфид, однако, оральные кинетосомы образуются de novo (Corliss, 1979; Foissner, 1996). При умножении кинетосом может сохраняться структура кинет (хапто- риды, простоматиды, циртофориды и кольподиды) или пролиферация идет так интенсивно, что исходная структура ресничных радон полностью исчезает (различные группы инфузорий). Наконец, у ряда инфузорий дифференцировка оральных структур связана с изменением топографии (ротацией или перемещением) зачатка. Завершение стоматогенеза обычно совпадает с началом цитокинеза — разделения материнской клетки надвое.
Согласно выше перечисленным особенностям, стоматогенез инфузорий подразделяют на следующие главные типы (Foissner, 1996): I — апокинетальный (ротовой аппарат развивается вне связи с предсущест- вующими структурами), II — паракинетальный (в связи со специальными посторальными соматическими кине- тами), III — буккокинетальный (в связи с предсущест- вующей оральной инфрацилиатурой), IV — телокине- тальный (в связи с предсуществующей соматической инфрацилиатурой) и V — миксокинетальный (когда в развитии принимают участие и соматические кинеты, и родительский ротовой аппарат). I тип стоматогенеза характерен для большинства представителей Oligothrichia и Hypotrichia; II — для многих Heterotrichea и Кагуоге- lictea; III — для большинства Oligohymenophorea; IV — для Protostomatea, Colpodea, Litostomatea и Phyllopha- ryngea и V — вероятно, для Apostomatia (Foissner, 1996).
Инцистирование. Весьма перспективным феноменом для изучения морфогенеза считаются процессы ин- и эксцистирования. Инцистирование — переход подвижных простейших в покоящуюся стадию, связанный с переживанием неблагоприятных условий или размножением, — явление широко распространенное среди инфузорий, хотя для некоторых групп (Paramecium) способности образовывать цисты не выявлено (Corliss, 1979; Фокин, 2002). Как было уже отмечено, существенную роль в образовании стенок цисты играет наличие у инфузорий муко- цист, за счет экструзии которых формируются ее слои. Очевидно, что процесс образования стенки цисты включает несколько стадий, так как обычно в ней выявляется несколько различно устроенных слоев (от 1 до 4) и зрелая циста часто представляет собой образование со сложной архитектоникой. При инцистировании большинство инфузорий теряет свой ротовой аппарат и соматическую цилиатуру (Matsusaka, Noguchi, 1989; Diaz et al., 2000). Образование цист деления или питания не приводит к глубоким изменениям в инцистирующейся клетке и сложность устройства самой цисты оказывается значительно меньшей.
Обратный процесс эксцистирования также связан с существеными морфогенетическими перестройками клетки инфузории, направленными на воссоздание активной, подвижной формы. При выходе из цисты покоя клетки либо прорывают ее стенку (Euplotes), либо покидают цисту через специальное отверстие (микропиле) — Nyctotherus, Bursaria.
ПОЛОВОЙ ПРОЦЕСС
Большую часть жизненного цикла инфузории не проявляют сексуальности, размножаясь бинарным делением (почкованием, стробиляцией, палинтомией). При этом с течением времени клетки клона обычно последовательно проходят ряд стадий: незрелости, взросления, зрелости и старости, в которые они могут (или не могут) вступать в половой процесс, имеющий у Ciliophora характер конъюгации. Собственно, возможность вступления в конъюгацию (или автогамию) связана именно с фазой зрелости и исследована прежде всего для нескольких модельных объектов (Tetrahymena, Paramecium, Euplotes, Blepharisma и некоторые другие) в лабораторных условиях. Однако для конъюгации необходимы не только определенный возраст клеток, но и принадлежность их к разным, но комплементарным типам спаривания, образующим сингены (Sonnebom, 1957; Dini, Nuberg, 1993; Miyake, 1996). Как правило, только в пределах одного сингена конъюгация инфузорий оказывается возможной и приводит к образованию жизнеспособного потомства. Число типов спаривания в одном сингене при бинарной системе всегда два, а при множественной — обычно кратно двум (Р. bursaria — 4, 6, 8; Uronychia — 10; Е. octocarinatus — 10, однако Tetrahymena — 7). Множественная система типов спаривания делится на го- мополярную и гетерополярную (Miyake, 1996). При первой каждый тип спаривания комплементарен всем остальным, а у видов со второй системой — некоторые комбинации оказываются некомплементарными. Последняя ситуация наиболее характерна для видов с большим числом типов спаривания (Euplotes, Stylonyc- hia). Типы спаривания у инфузорий (в зависимости от видовой принадлежности) определяются генотипически или эпигенетически и иногда (Р. multimicronuc- leatum, синген 2) изменяются во времени (Barnett, 1966).Число сингенов (биологических видов) существенно варьирует у представителей Ciliophora. Так, среди парамеций встречаются виды, для которых известно от одного сингена (Р. polycaryum) до 15—16 (комплекс Р. aurelia, Р. caudatum). Инфузорий с бинарной системой типов спаривания до сих пор известно крайне мало: виды Paramecium (за исключением Р. putrinum и Р. bursaria), Aspidisca, Blepharisma, Glaucoma, E. Crassus, Tokophrya, в то время как остальные изученные в этом отношении инфузории обладают множественной системой (Dini, Nuberg, 1993;Miyake, 1996; Fokin etal., 2001). Для значительного числа видов Ciliophora система типов спаривания до сих пор не установлена.
Возможность вступления клеток в конъюгацию определяется окончанием периода незрелости, который может быть продолжительным — несколько месяцев (Р. bursaria)', умеренным — около месяца (Р. caudatum); коротким — 1—3 нед (Р. jenningsi), или может отсутствовать вообще (Р. tetraurelia).
На основе взаимораспознавания партнеров происходит формирование конъюгационной пары, клетки в которой претерпевают полный или частичный процесс конъюгации. Иногда партнеры расходятся раньше стадии обмена пронуклеусами, и тогда ядерная реорганизация в клетках может проходить по типу автогамии. Насколько часто конъюгация происходит в природных условиях, для большинства видов неизвестно, а для некоторых инфузорий конъюгация вообще никогда не наблюдалась (Lucchesi, Santangelo, 2004).
В большинстве случаев у инфузорий в конъюгациювступают клетки одинакового размера, которые образуют две зиготы (пару эксконъюгантов). Такая конъюгация называется изоморфной бизиготической. Каждый конъюгант дает два (иногда более) гаплоидных ядра, одно из которых стационарное, а второе — мигрирующее. Реципрокное слияние пронуклеусов (в результате перехода мигрирующих ядер в конъюгационного партнера) приводит к образованию двух диплоидных зигот (синкарионов).
У Peritrichia и некоторых Suctoria конъюгируют клетки, значительно различающиеся по размерам (макро- и микроконъюганты), и в результате процесса ядерной реорганизации в конъюгантах образуется по одному про нуклеусу. Меньший конъюгант сливается с более крупным, образуя лишь одну зиготу (один эксконъюгант). Такой вариант конъюгации называется анизоморфной монозиготической. В ряде случаев (Ephelota, Metopus, Urostyla, Chonotricha) обнаружена также изоморфная монозиготическая конъюгация (Янковский, 1973; Miyake, 1996). Помимо этого, при определенных условиях возможны конъюгация между клетками одного клона (селфинг) или половой процесс в пределах единственной клетки (автогамия): Р. aurelia комплекс, Р. jenningsi, Р. polycaryum, Е. minuta, Т. rostrata (Wichterman, 1986; Miyake, 1996).
В основе взаимораспознавания конъюгантов может лежать прямое или непрямое взаимодействие между клетками. Так, инфузории Paramecium и Tetrahymena должны вступать в кратковременный механический контакт (по-видимому, столкновения при плавании) перед образованием пар. В этот период костимуляции происходит модификация части кортекса: у тетрахимен — переднего конца клетки, у парамеций — части вентральной поверхности, окружающей буккальную зону (Fujishima, 1988; Miyake, 1996). Этот этап может занимать несколько часов. У других инфузорий (В1е- phrisma, Euplotes, Dileptus) предконъюгационное взаимодействие между партнерами непрямое: посредством растворимых в воде гамонов и соответствующей системы рецепторов (Luporini, Miceli, 1986; Miyake, 1996).
Процесс конъюгации распадается на прогамную и метагамную фазы. Прогамная фаза начинается с объединения партнеров и заканчивается образованием зиготы — синкариона; ме- тагамная — простирается от момента образования синкариона до восстановления вегетативного ядер- ного аппарата. Часть продуктов деления микронуклеуса на прогамной фазе (всегда) и на метагамной фазе (часто) дегенерирует, что приводит к восстановлению исходного ядерного аппарата обычно в ходе двух или трех постконъюгационных делений эксконъ- югантов (Miyake, 1996; Fokin et al., 2001). Конъюгаци- онный цикл считается завершенным при вступлении потомков эксконъюгантов в следующий половой процесс.
Старое соматическое ядро у эксконъюгантов дегенерирует, однако время и особенности этого процесса значительно различаются в разных группах инфузорий, часто даже близкородственных. Весьма характерным изменением старого макронуклеуса в ходе конъюгации является его фрагментация, которая может начинаться как на прогамной, так и на метагамной (наиболее часто) фазе процесса (Raikov, 1972; Fokin et al., 2001). Нередко старый макронуклеус дегенерирует без изменения (Karyorelictea, многие Heterotri- chea, Tetrahymena, Р. bursaria), а иногда началу этого процесса предшествует дополнительное деление макронуклеуса (Urocentrum, некоторые Euplotes, Р. dubosequi).
Помимо обычной конъюгации для инфузорий (Paramecium) известна цитогамия, при которой в образовавшийся паре конъюгантов ядерная реорганизация проходит без обмена пронуклеусами, что, следовательно, можно рассматривать как парную автогамию (Wichterman, 1986). «Полусексуальная» конъюгация была обнаружена у Bl. japonicum (Miyake et al., 1991). У этого мультимикронуклеарного вида часть микронуклеусов не вступает в мейоз, а прямо дифференцируется в макронуклеусы в начале конъюгационного процесса. Остальные микронуклеусы претерпевают нормальную ядерную реорганизацию с образованием новых элементов ядерного аппарата. Таким образом, в одной клетке (в результате доминирования одной из групп макронуклеусов) могут быть либо все ядра, возникшие половым путем, либо только микронуклеусы.
Как уже было сказано, для некоторых инфузорий (иногда только для некоторых линий одного вида — Е. minuta, Е. crassus) помимо конъюгации известна автогамия, при которой процессы ядерной реорганизации проходят практически по такой же схеме, но в пределах одной клетки (Nobili, 1966; Miyake, 1996). Среди комплекса видов Tetrahymena у Т. rostrata автогамия происходит в цистах (Corliss, 1965).
СИМБИОЗ
Долгое время наличие симбиотических организмов (в широком смысле этого понятия, предложенного Де Бари (De Вагу, 1879)) в составе или на поверхности клетки Protista воспринималось как ситуация случайная и нестабильная (Kirby, 1941), хотя еще в XIX—начале XX в. некоторые отечественные протистологи правильно оценили возможное значение этого феномена (Ценковский, 1856; Фаминцын, 1889; Мережковский, 1909). С усовершенствованием микроскопической техники и особенно с появлением электронного микроскопа стало очевидно, что в большинстве групп простейших отсутствие симбионтов, особенно бактериальных, является скорее исключением. Именно к таким таксонам относится и тип Ciliophora (Preer, Preer, 1984; Heckmann, Gortz, 1991; Фокин, 1993a; Fokin, 2004a).Симбиоз у инфузорий дает примеры разнообразных комбинаций этих простейших с про- и (иногда) эукариотами. Очевидно, некоторые особенности Ciliophora (например, почти исключительно распространенный у них фаготрофный тип питания) создают предпосылки для возникновения этого явления. Характер взаимодействия между партнерами в симбиотических системах Ciliophora далеко не всегда подробно исследовался, но, очевидно, он может представлять все переходы от паразитизма до мутуализма (Gortz, 1996).
В последние время, в том числе и в связи с развитием симбиотической теории происхождения эука- риотной клетки, симбиозу у инфузорий, особенно внутриклеточному, уделялось значительное внимание. Адаптации и взаимоотношения партнеров в симбиотической системе, экологическая и эволюционная значимость симбиоза у инфузорий, механизмы инфекции — вот только часть вопросов, интенсивно изучаемых ци- лиатологами (Gortz, 1988, 1996). При этом сами инфузории также могут выступать в роли симбионтов (комменсалов, паразитов) различных многоклеточных и изредка одноклеточных животных.
Бактериальные эндобионты могут заселять все основные компартменты клетки инфузорий: они встречаются в кортексе, эндоплазме, макро- и микронуклеусах и перенуклеарном пространстве, и даже в митохондриях. Для клетки хозяина, за некоторым исключением (часть инфузорий анаэробных местообитаний, несколько видов Euplotes и, возможно, Climacostomum), такое сожительство факультативно и, как правило, лишь незначительная доля популяции заражена бактериями, хотя в некоторых из них зараженными оказываются почти все клетки (Lloyd et al., 1996). Следует, однако, оговориться, так как в большинстве случаев симбиоз инфузорий с бактериями глубоко не исследовался, и сказать, облигатный ли он или факультативный (для хозяина), иногда нельзя. С другой стороны, подавляющее большинство эндоби- онтных бактерий, обнаруженных в инфузориях, не может существовать вне хозяина, и, следовательно, они являются облигатными симбионтами. Поддерживать эти микроорганизмы вне тела инфузории обычно не удается даже на весьма сложных искусственных средах (Soldo, 1973, 1974; Gortz, 1983). Исключение составляют метаногенные бактерии, обычные в цитоплазме анаэробных инфузорий: по крайней мере, представители родов Methanobacterium и Methanoplanus могут культивироватся вне тела хозяина (Bruggen et al., 1984, 1986).
Существенный интерес представляет группа бактерий, разнородных (по-видимому) в филогенетическом отношении, которые сообщают хозяину killer-свойства (способность убивать инфузорий, не содержащих этих эндоцитобионтов). По преимуществу это представители рода Caedibacter, наиболее исследованные для комплекса видов Р. aurelia (Preer et al., 1974; Quackenbush, 1988), хотя они известны также для Р. caudatum, Р. dubosequi и для Spirostomum (Schmidt et al., 1987; Fokin, Gortz, 1993; Fokin et al., 2003). Характерной чертой этих бактерий считается присутствие у них R-тел — скрученных в виде цилиндра протеиназных лент длиной до 20 мкм и размерами (в упакованном виде) — 0.4—0.5 X 0.4—0.8 мкм. С этими структурами обычно связаны капсиды бактериофагов и способность вызывать killer-эффект посредством токсина неизвестной, но, по-видимому, белковой природы.
Эффект может быть дистанционным, что предполагает выход токсина (или структур, его содержащих) во внешнюю среду, или контактным, когда для проявления killer-эффекта клетка, несущая бактерии, должна вступить в конъюгацию с чувствительной парамецией. Другие бактерии, не образующие R-тел, сообщают killer-свойства Parauronema acutum (Soldo, 1973) иР. nephridiatum (Фокин, 1989), причем в последнем случае эффект наступает только при конъюгации с апосимби- онтным партнером — mate-killer (рис. 258). Очевидно, что такие симбионты дают своему хозяину определенные преимущества при внутривидовой, а иногда и межвидовой борьбе за источники пищи. Экологическое значение такого рода ассоциаций для клетки-хозяина было экспериментально продемонстрировано в натурных экспериментах (Landis, 1988).
Длительное время в качестве модельной для изучения взаимодействий про- и эукариотических партнеров по симбиозу используется система Parameci- w/н-бактерии рода Holospora (Осипов, 1981; Gortz, 1983а, 1988, 1996; Фокин, 1993; Сабанеева и др., 2002; Fokin, 2004а). Облигатные, высоко инфекционные бактериальные эндонуклеобионты парамеций, относимые к роду Holospora, обладают жизненным циклом, состоящим из инфекционной и вегетативной фаз. В клетку хозяина в результате нормального процесса фагоцитоза могут попадать обе формы этих бактерий, однако только инфекционные, имеющие специальную клеточную дифференцировку (наличие цитоплазматической, периплазматической части и переднего «акросомного» конца) и размеры от 10 до 20 мкм, служат источником заражения новых хозяев. После слияния первичной фагосомы с ацидосомами инфекционные формы холоспор «активируются» и покидают пищеварительную вакуоль хозяина в окружении части ее мембраны. Далее, в окружении мембраного комплекса, возникающего за счет клетки-хозяина, активированные инфекционные формы холоспор, оказавшиеся возле ядер, взаимодействуют (согласно их ядерной специфичности) с оболочкой макро- или микронуклеуса парамеции. В результате этого взаимодействия они оказываются включенными в кариоплазму, где через непродолжительное время фрагментируются, переходя к вегетативной части своего жизненного цикла. Процесс инфекции занимает не более 1 ч. Жизненный цикл бактерий замыкается, когда после нескольких дней вегетации в ядрах инфузории дифференцируются новые инфекционные формы, которые выводятся из клеткй хозяина во внешнюю среду и заражают новых хозяев-парамеций.
Значительный набор этих бактерий (10 видов, инфицирующих 6 видов парамеций), возможность лабораторного заражения холоспорами апосимбиотиче- ских клеток парамеций и экспериментального конструирования сочетаний, в которых эндобионты и хозяева имеют различные морфологические и генетические особенности, делают эту симбиотическую систему очень удобной для решения ряда важных вопросов симбионтологии и клеточной биологии. Одними из центральных в этом ряду являются проблема клеточного распознавания и вопросы, связанные с путями инфекции прокариотами эукариотической клетки (Осипов, 1981; Gortz, 1988, 1996).
В сравнении со жгутиконосцами (Ball, 1969) инфузории имеют не так много бактериальных эктобионтов (эпибионтов). Весьма распространены эпибионтные бактерии, как уже отмечалось, у многих инфузорий, живущих в условиях дефицита кислорода. Вероятно, для некоторых из них симбиоз с бактериями облигатен. Среди примеров такого рода симбиоза такие интересные и впечатляющие случаи, как облигатный симбиоз карио- реликтиды К. latum и перитрихи Z. niveum с серными бактериями (Raikov, 1974; Fenchel, Finlay, 1989; Bau- er-Nebelsick et al., 1996). Если для Kentrophoros бактерии — это источник питания, то роль прокариот в жизни Zoothamnium пока не была детально исследована. Бактериальная «шуба», плотно покрывающая всю поверхность микро- и макрозооидов и переносимая последними на новые колонии — аргумент в пользу взаимовыгодного такого сожительства.
Интересный пример облигатного симбиоза демонстрирует морская wmmymxz Euplotidium itoi. Эпибион- ты широкой полосой покрывают часть маргинальной поверхности клетки, располагаясь в индивидуальных углублениях кортекса хозяина (Vemi, Rosati, 1990; Rosati, 1999). Ранее они рассматривались как специализированные экструсомы, поскольку эти микроорганизмы имеют сложный аппарат экструзии, несколько напоминающий R-тела бактерий рода Caedibacter и аппарат экструзии криптомонад (Gortz, 1996). Действительно, при определенных воздействиях происходит «выстреливание» содержимого аппарата, хотя его значение пока непонятно. Одно из возможных объяснений сводится к тому, что эти микроорганизмы действительно функционально замещают защитные экструсомы, которыми Euplotidium не обладает (Rosati et al., 1999).
Одним из интенсивно изучаемых примеров симбиотической системы между инфузориями и водорослями служит Р. bursaria, практически всегда имеющая в своей цитоплазме 200—300 зеленых водорослей рода Chlorella (Reisser, et al., 1985; Reisser, 1986). Водоросли располагаются в специальных (периальгальных) вакуолях, образованных мембранами хозяина и не способных сливаться с лизосомами, что предохраняет симбионтов от переваривания. Находясь в цитоплазме парамеций, водоросли также оказываются защищенными от вирусов, часто лизирующих несимбиотические хлореллы или водоросли, покинувшие клетку-хозяина (Reisser et al., 1988).
Парамеции, содержащие водоросли, в свою очередь оказываются в лучшем положении, чем апосимбионтные клетки при недостатке пищи: некоторые «зеленые» инфузории могут существовать при достаточной освещенности без дополнительного питания месяцами. Клетки Chlorella продуцируют значительное количество мальтозы, которая используется хозяином- парамецией. Такая ситуация меняет поведение инфузорий, которые предпочитают в противоположность своим апосимбиотическим родственникам более освещенные местообитания (Cronkite, 1986). Наличие хлорелл в цитоплазме Р. bursaria делает хозяина значительно более устойчивым к всевозможным стрессам, в том числе к бактериальной инфекции (Gortz, 1983b). Кроме того, показано, что «зеленые» парамеции реже служат объектом нападения D. nasutum, что говорит о наличии защитной функции хлорелл в этой симбиотической системе (Berger, 1980).
Хотя большинство Ciliophora — свободноживущие простейшие, среди них встречаются и симбиотические виды (Gortz, 1996; Bradbury, 1996). К ним относятся экто- и эндокомменсалы различных беспозвоночных и позвоночных животных и незначительное число паразитических инфузорий. Так, представители Ophryoscolecidae (Litostomatea) во множестве населяют рубец жвачных; виды Nycthoterus (Armophorea) — кишечный тракт бесхвостых амфибий; инфузории группы Phylasterida (Oligohymenophorea) — живут на кишечнополостных, заселяют мантийную полость разнообразных моллюсков, кишечный тракт иглокожих и рыб; представители Ophryoglenina (Oligohymenophorea) паразитируют на рыбах (Ichthyophthirius), в моллюске Dreissena и поденке Caenis (Ophryoglena). К немногим патогенным инфузориям относится и единственный вид, заражающий человека, — Balantidium (Litostomatea). Нередко к эндопаразитизму переходят представители Tetrahymena (Oligohymenophorea): они обнаружены в личинках мошки, москитов и даже в мочеполовой системе млекопитающих (Lynn et al., 2000). Некоторые Suctoria (Sphaerophrya) паразитируют в клетке инфузорий: Euplotes, Uroleptus, Urostyla, Bursaria, Paramecium, и в то же время Cyathodinium (Suctoria, Evaginogenida) заселяют пищеварительный тракт гвинейских свиней, a Allantosoma multisuctores (Suctoria, Endogenida) была обнаружена в африканском носороге (Hoven et al., 1998).
Ряд эпибионтных инфузорий известен удивительной хозяинной и топографической специфичностью — они поселяются в строго определенных местах покровов хозяина (прежде всего различных членистоногих). Так, Dendrocometesparadoxus встречается только на жабрах бокоплава Gammarus pulex. На G. locustra поселяется около 10 различных видов эпибионтов, весьма определенным образом распределяясь по членистым придаткам этого ракообразного (Fenchel, 1965). Такая же тенденция строгой приуроченности к участкам тела рачка наблюдается у хонотрих, причем обычно инфузории прикрепляются к придаткам, находящимся в постоянном движении, что создает оптимальные условия для их питания (Янковский, 1973).
Для неподвижных инфузорий-эпибионтов большое значение имеет плотность популяции хозяина, что значительно облегчает их расселение. Некоторые же эпи- биопты не закрепляются постоянно на одном хозяине, а могут менять субстрат, активно плавая. В таких случаях часто вырабатываются специальные прикрепительные приспособления типа диска у Trichodina. Представители этого рода перитрих из отряда Mobilida (Oligohymenophorea) встречаются на гидрах, в разнообразных моллюсках, у гребневиков (Esters et al., 1997). Многие виды паразитируют на рыбах и даже на амфибиях. Kerona polyporum (Spirotrichea) в противоположность триходинам не выработала специальный прикрепительный аппарат, но за счет углубления на вентральной поверхности клетки может прочно присасываться к покровам хозяина-гидры.
Эпибионтные Suctoria (Phyllopharyngea) в качестве хозяев используют широкий набор беспозвоночных животных, в том числе и инфузорий. Наиболее обычны эти эпибионты (паразиты) на ротовых частях поли- хет, на гидрополипах и ракообразных. Из инфузорий они предпочитают поселяться на Peritrichia (Oligohymenophorea).
ЖИЗНЕННЫЕ ЦИКЛЫ И ПОЛИМОРФИЗМ
В большинстве случаев жизненный цикл у инфузорий состоит из значительного числа бесполых поколений, когда клетка (трофонт) размножается поперечным делением, давая фенотипически и генотипически себе подобных, и периода конъюгации, в результате которой образуются генотипически новые особи, чем жизненный цикл и завершается. Наличие диморфных или полиморфных стадий, процесса почкования и ин- цистирования в жизненном цикле многих инфузорий делает его иногда достаточно сложным. Как правило, сложными жизненными циклами обладают сидячие, комменсальные и редкие среди инфузорий паразитические формы, что связано с их размножением и распространением (поисками и сменой субстратов или хозяев).В качестве максимально простого жизненного цикла можно привести таковой парамеций, для которых неизвестна стадия цистообразования и которые не дают при делении полиморфных форм (Wichterman, 1986). Большое число инфузорий образуют цисты покоя (прежде всего при недостатке питания или другой форме стресса), а некоторые — и цисты размножения, что приводит к появлению известного полиморфизма в пределах одного жизненного цикла. Полиморфизм может возникать и в результате изменения условий окружающей среды (прежде всего соотношения пищи и хищников). Так, у Т. vorax при обилии мелких инфузорий, служащих ей пищей, появляется мак- ростомная форма инфузории, способная питаться объектами, сравнимыми с ней по величине (Williams, 1961). Сходное явление было обнаружено для Е. versa- tilis (Tuffrau et al., 2000), у которого при обилии пищи(бактерий) деление клетки происходит неравномерно и протер оказывается меньше, чем опистер. Возникшие гигантские клетки (увеличение размеров может быть более чем в 2 раза) становятся каннибалами.
Наличие в среде значительного числа хищников также вызывает появление гигантских форм Euplotes, которые за счет размеров и формы своего тела становятся недоступными в качестве пищевого объекта (Tuffrau, 1959; Kuhlmann, Heckmann, 1987; Kusch, 1993).
Другим примером диморфизма могут служить клеточные вариации в жизненных циклах сидячих инфузорий (хонотрих, перитрих и сукторий), где «личиночные» формы — «бродяжки» — обычно мелкие подвижные клетки, служащие для расселения, тогда как «взрослые» инфузории значительно крупнее, лишены подвижности и соматической (или всей) цилиатуры. Для некоторых колониальных инфузорий в пределах колонии также отмечен полиморфизм зооидов (Bauer-Nebelsick et al., 1996). Для представителей этих же групп, а также некоторых гименостоматид и астоматид характерны почкование (моно- и полигем- мия) и палинтомия.
Так, для Chonotricha жизненный цикл включает рост и питание трофонтов (активно питающихся и растущих форм), образование томонтов (не- питающихся делящихся или почкующихся форм), которые продуцируют томиты (почки — до отделения от материнской клетки и бродяжки — после отделения). Томиты не питаются, оседают на субстрате и начинают метаморфоз, превращаясь в трофонтов (Янковский, 1973). Не все трофонты вырастают во взрослые формы; начиная со стадии молодого трофонта развитие может пойти двумя путями — с конъюгацией или без нее. К конъюгации, по-видимому, способны только молодые трофонты, и этот процесс коррелирует с линькой хозяина (рачка). Молодые эксконъюганты (трофонты) возобновляют питание и рост и, достигнув максимальной величины, приступают к наружному или внутреннему почкованию (Янковский, 1973).
Известный полиморфизм в жизненном цикле демонстрируют и сидячие инфузории, обладающие лорикой. Так, Ей. uhligi в результате деления дает типичную форму (трофонта), помещенную в лорику, и подвижную бродяжку, которая, закрепившись на субстрате, развивает из зачатка характерный ротовой аппарат и секретирует лорику de novo (Mulisch, Patterson, 1987).
Исключительное явление среди инфузорий представляет собой жизненный цикл колькодиды Sorogena stoianovitchae (Olive, Blanton, 1980), образующей воздушный сорокарп, весьма напоминающий сходные образования у представителей Mycetozoa. Сорогена питается мелкими представителями рода Colpoda. При уменьшении количества пищи клетки Сорочены образуют агрегат на поверхности воды, оседая на бактериальную пленку. Из этого агрегата вырастает воздушное плодовое тело (сорокарп), несущее множество со- роцист. После нескольких дней свободного плавания зрелый спорокарп разрушается и цисты падают на поверхность воды (в сухом состоянии они сохраняют жизнеспособность около двух лет), где через пору из них выходят активно плавающие клетки.
Достаточно распространены сложные жизненные циклы у паразитических инфузорий, относящихся к подклассам Hymenostomatia и Apostomatia (Oligohy- menophorea): Ichthyophthirius, Cryptocaryon и Vampy- rophrya (Bradbury, 1996). Представители первого рода паразитических инфузорий редко встречаются в природе, но являются серьезной проблемой для прудовых и озерных рыборазводных предприятий и аквариумов. Инфекционные формы Ichthyophthirius (мелкие терон- ты, имеющие развитую цилиатуру и Либеркюнову органеллу — характерный признак всех Ophryoglenina), тысячами выходят из цист размножения, которые находятся на дне водоема, и заражают хозяина (различных рыб). В эпителии (кожа и жабры) хозяина инфузории превращаются в активно питающихся трофонтов, достигающих размеров более 1 мм, лишенных органеллы Либеркюна и имеющих измененную цилиатуру (Ewing, Косап, 1986). После нескольких дней питания трофонты покидают хозяина и, опустившись на дно, инцистируются (Ewing, Косап, 1987). В цистах размножения в результате палинтомии образуется огромное количество мелких томитов, которые при благоприятных условиях (значительном количестве рыб подходящих видов) замыкают жизненный цикл этой инфузории (Bradbury, 1996).
Апостоматида Vampyrophrya — эктопаразит пелагических копепод (Grimes, Bradbury, 1992). Томиты этой инфузории находят хозяина и прикрепляются на поверхности его хитинового покрова. В случае повреждения панциря вампирофрии быстро эксцистиру- ются и переходят к питанию тканями хозяина, после чего инцистируются и в цисте размножения образуют множество томитов. Последние покидают цисту и, активно плавая, ищут новых рачков, на покровах которых оседают. Если зараженные рачки поедаются другими животными (некоторыми Cnidaria или Chae- tognata), инфузории могут эксцистироваться в кишке хищника и питаться за его счет, давая более мелких трофонтов (Bradbury, 1996).
ФИЛОГЕНИЯ И ЭВОЛЮЦИЯ
По современным представлениям инфузории произошли от фаготрофных бентических жгутиконосцев, обладавших дикинетидами с фибриллярными дериватами, среди которых, по-видимому, наибольшее развитие имели посцилиарные микротрубочки. В результате полимеризации дикинетид предков образовалась пароральная цилиатура инфузорий, позднее давшая начало адоральным мембранеллам и соматической цилиатуре (Bardele, 1992; Schlegel, Eisler, 1996).Сравнительный анализ ультраструктурных особенностей (наличие альвеол) и последовательностей нуклеотидов в генах рРНК показывает, что из ныне существующих групп наиболее близки к инфузориям Dino- flagellata и Apicomplexa (Corliss, 1988; Schlegel, 1991; Wolters, 1991; Wright et al., 1997; Kuvardinaet al., 2002), образующие в системе Кавалье-Смита инфрацарство Alveolata (Corliss, 1998; Patterson, 2000). Отсутствие у динофитовых гистонов и присутствие конденсированных хромосом и внеядерного митотического аппарата давали основания рассматривать эту группу как предковую для эукариот. Молекулярно-биологические данные, однако, указывают, скорее, на происхождение современных Dinoflagellata в период возникновения большинства других групп: животных, растений, грибов, красных водорослей (Sogin et al., 1986).
Палеонтологические данные об инфузориях ограничены находками некоторых тинтинид, обладавших лорикой (Ордовик, Палеозойская эра, 400—450 млн лет назад) и фолликулин. По морфологическим особенностям обнаруженных окаменелостей эти инфузории мало отличаются от современной фауны и могут быть помещены в соответствующие современные семейства и даже роды (Corliss, 1979). Отмечены также находки гипотрих, нассулид, кольподид и пеникулин в янтаре (возрастом 220—230 млн лет) из триаса. Причем в последнем случае находка была описана как новый (вымерший) вид: Р. triassicum (Schonbom et al., 1999). Очевидно, что инфузории появились значительно раньше — в палеозое (Wright, Lynn, 1997а).
Инфузории — эволюционно очень продвинутая группа протистов. Ее представители обладают значительным числом молекулярных и морфологических особенностей. С точки зрения общего плана строения (уникальных черт) — ресничного покрова с эктоплазматической фибриллярной системой и гетероморфного ядерного аппарата — Ciliophora считается тем не менее наиболее гомогенным типом среди прочих Protista. Частные планы строения групп, в него входящих, их цитоархитектоника, обладающая множественными модификациями, и прочие особенности позволяют анализировать эволюционные (филогенетические) взаимоотношения в пределах типа, основываясь на морфологических и биологических данных (Bardele, 1981, 1989; Small, Lynn, 1981, 1985; Puytorac et al., 1984, 1994; Герасимова, 1989; Lynn, 2001, 2003).
Изначально это были только сведения относительно характера соматической и оральной цилиатуры. Значительную помощь систематикам на этом этапе оказало появление методик импрегнации кинетома солями серебра (Klein, 1926; Chatton, Lwoff, 1930; Gelei, Horvath, 1931). Применение электронного микроскопа сместило «центр тяжести» на данные по ультраструктуре и прежде всего инфрацилиатуре инфузорий (Puytorac et al., 1974; Lynn, 1975, 1976, 1981; Герасимова, Серавин, 1976; Серавин, Герасимова, 1977; Small, Lynn, 1981). Однако наиболее известная классификация инфузорий 70-х гг. (Corliss, 1974, 1979), используемая в учебниках до сегодняшнего дня, во многом еще была ориентирована на данные по буккальному и общему морфогенезу различных инфузорий (Lynn, 1996b). Существенное значение для ее появления сыграли воззрения на систему инфузорий Янковского (1967), а информация по ультраструктурной организации Ciliophora еще не использовалась в этой системе в полной мере (Corliss, 1979). Однако вместо двух подклассов, Holotricha и Spirotricha, на которые были разделены представители класса Ciliata, подтипа Ciliophora (Corliss, 1961), в системе 1979 г. удалось обосновать 3 класса типа Ciliophora, которые включали уже 7 подклассов и 23 отряда.
Исследование кольподид, предпринятое Лином (Lynn, 1975), показало, что в этой группе разнообразие организации оральных структур весьма значительно, в то время как соматическая цилиатура устроена однообразно. Очевидно, Герасимовой и Серавиным, с одной стороны, и Лином — с другой, практически одновременно и независимо была высказана идея о консервативности организации соматических кинетосом и их дериватов у инфузорий и возможности построения на основе этого признака филогенетической схемы классификации Ciliophora (Герасимова, Серевин, 1976; Lynn, 1976). В результате анализа ультраструктуры соматической цилиатуры у представителей основных групп инфузорий в 1981 г. Смолом и Лином была предложена макросистема Ciliophora, где инфузории были разбиты на 3 подтипа, 8 классов, 15 подклассов и 48 отрядов, прежде всего по данным электронной микроскопии (Small, Lynn, 1981, 1985). Такой подход позволил определить место в системе для ряда родов, положение которых прежде было проблематично или неверно (например, для Bursaria и Sorogena), и обосновать классы Ciliophora как монофилетические образования.
В основу построения системы Ciliophora Корлисом была положена организация прежде всего орального аппарата: Kinetofragminophora (с простым ротовым аппаратом), более продвинутые Oligohymenophora (с усложненным ротовым аппаратом) и Polyhymenophora (с наиболее сложным и хорошо развитым ротовым аппаратом), что отвечало, по воззрениям большинства протозоологов, филогенетической линии развития Ciliophora. Схема, построенная на основе структурного консерватизма соматического кортекса (Small, Lynn, 1985; Lynn, Corliss, 1991), радикально отличалась от предыдущей. Kinetofragminophora и Polyhymenophora перестали существовать, оказавшись полифилити- ческими группировками, и только Oligohymenophorea сохранили статус класса. При этом подкласс Spirotri- cha получил ранг класса, так же как и отряды (иногда в несколько измененном составе) Karyorelictida, Prosto- matida, Colpodida, Nassulida, (Small, Lynn, 1985).
Начиная с середины 80-х гг. XX в. в протистологии явно начали преобладать использование в филогенетических построениях молекулярных маркеров: энзимов, ДНК/ДНК гибридизации и во все большей степени сравнения нуклеотидных последовательностей гена рРНК (Schlegel, Eisler,1996). Аналогичным образом развивалась и филогенетика инфузорий (Lynn, 1996а, 1996b; Schlegel, Eisler, 1996; Lynn, Small, 1997; Struder-Kypke et al., 2000; Miao, Shen, 2001; Wright, Co- lomi, 2002; Lynn, 2003).
Основным результатом, сразу полученным при использовании сравнительного анализа последовательностей нуклеотидов в гене рРНК, оказалась диаметрально противоположная топология примитивных (согласно прежним воззрениям) и продвинутых групп Ciliophora (Greenwood et al., 1991; Schlegel, 1991; Bar- dele, 1992). Так, Bl. americanum (Polyhymenophora), которую помещали рядом с гипотрихами, согласно анализу рРНК, дивергировала в процессе эволюции Ciliophora очень рано вместе с кариореликтидами, так же как и Stentor (Greenwood et al., 1991; Schlegel, 1991), что подтвердило выделение класса Heterotrichea, сделанное по морфологическим признакам. В то же время Prostomatea, считавшаяся одной из примитивных групп инфузорий (Puytorac et al., 1987; Bardele, 1989), оказалась близка к Colpodea, Plagiopylea и Oligohymenophorea(Fleury et al., 1992; Shin, et al., 2000; Striider- Kypke et al., 2000; Wright, Collomi, 2002).
Сравнительно-морфологические (морфогенетические) данные дают основание думать, что Prostomatea возникли в результате апикального смещения ротового комплекса и его редукции с последующей утратой адоральной цилиатуры, в то время как парораль стала циркуморальной цилиатурой. Видимо, нечто похожее произошло с гапторидами, трихостоматидами и энто- диниоморфидами (Litostomatea), у которых утраченная первичная ротовая цилиатура была замещена за счет соматической (Bardele, 1992; Foissner, 1996; Lynn, 2001). Филогенетическая схема инфузорий, построенная прежде всего на базе онтогенетических данных (Foissner, 1996), во многих деталях совпала с данными молекулярного анализа.
Дополнение дендрограммы, построенной на основании анализа рРНК, данными по новым видам (первоначально в анализ были вовлечены в основном представители Oligohymenophora и Polyhymenophora), позволило к настоящему времени получить репрезентативные схемы филогенетических связей основных классов Ciliophora (Leipe et al., 1994; Hammerschmidt et al., 1996; Wright, Lynn, 1997b; Striider-Kypke et al., 2000; Shin et al., 2000; Baumgartner et al., 2002; Lynn, 2003). Уже полностью или частично известны нуклеотидные последовательности гена рРНК более чем для 120 видов инфузорий. Тем не менее для некоторых групп этих простейших реконструкция филогенеза с использованием данных молекулярной биологии до сих пор отсутствует или полученные картины крайне фрагментарны и нестабильны из-за использования для анализа лишь единичных видов. Очевидно, что для уточнения и стабилизации позиций отдельных групп потребуется дополнительное исследование видов, до сих пор нс вовлеченных в анализ.
Молекулярные данные подтвердили монофилию Ciliophora и существование теперь уже 10 (Plagiopylia и Spirotrichia переведены в ранг классов) классов этих простейших (Lynn, Small, 1997, 2000; Shin et al., 2000; Baumgartner et al., 2002; Wright, Colorni, 2002). Очевидно, что статус класса заслуживает и дивергентная группа Armophorca: отряд Armophorida (Metopidae, Caenomorphidac) 4- отряд Clevelandellida (Nyctotheridae, Clevelandellidae, Sicuophoridae) + отряд Odontostomati- da, хотя ее положение в системе пока неоднозначно (Wright, Lynn, 1997b; Shin et al., 2000; Baumgartner et al., 2002; Wright, Colorni, 2002; Lynn, Striider-Kypke, 2002; Lynn, 2003).
Разделение тина на Postciliodesmatophora, Rhabdo- phora и Cyrtophora (первый подтип был выделен по преобладанию среди дериватов кинетосом посцилиар- ных микротрубочек, а в торые два — по организации ротового аппарата) (Small, Lynn, 1985) уступило теперь место двум подтипам. В качестве дополнительного признака при этом было использовано положение микротрубочек при делении макронуклеуса (Lynn, 1996b). Сохранивший свое название подтип Postciliodesmatophora включает наиболее древних инфузорий: Кагуоге- lictea и Heterotrichea. У представителей этих классов микротрубочки при делении соматического ядра располагаются в цитоплазме. Все остальные инфузории пока объединены в подтип Intramacronucleata (с внутримакронуклсарными микротрубочками), хотя автор системы признает возможность находки более фундаментальных признаков, отличающих эти группы (Lynn, 1996b). Наиболее филогенетически близкими к Helcrotrichca могут считаться Spirotrichea и Litostoma- tea (Lcipc cl al., 1994), хотя значительная бифуркация на дендрограммах в подтипе Intramacronucleata делает такие выводы предварительными.
Взаимоотношения в классе Spirotrichea достаточно полно проанализированы в последнее время молекулярными методами (Shin et al., 2000; Chen, Song, 2002). В результате обоснован новый подкласс Phacodiniidia и подтверждено существование подкласса Protocruzii- dia (Puytorac et al., 1984). Оба подкласса, так же как Choreotrichia и Oligotrichia, признаны по данным молекулярного анализа монофилетическими. Анализ филогенетических связей в подклассе Hypotrichia (Chen, Song, 2002) показал, что эта группа также монофилетическая, с базальной позицией рода Euplotes.
Монофилия Prostomatea, Colpodea и Oligohymeno- phorea также получила подтверждение в работах последних лет (Foissner, 1996; Striider-Kypke et al., 2000; Lynn, 2001). Очевидно, что эти классы достаточно тесно связаны между собой. Несомненна близость к этой группировке и Plagiopylea (Baumgartner et al., 2002), обладающих рядом «олигоменофорных» черт. По данным Райта с соавт. (Wright et al., 1997), плагиопилиды должны даже составить подкласс Plagiopylia в составе Oligohymenophorea, что, правда, не подтверждается более поздними данными (Striider-Kypke et al., 2000; Wright, Colomi, 2002). С другой стороны, показано близкое родство между Colpodea и Nassophorea (Bern- hard et al., 1995; Striider-Kypke et al., 2000).
Полномасштабный анализ филогенетических связей внутри одного из крупнейших классов Ciliophora — Oligohymenophorea пока затруднен из-за отсутствия данных по двум подклассам: Astomatia и Aposto- matia. Сделанные работы (Striider-Kypke et al., 2000; Miao et al. 2001; рис. 266) позволили, однако, представить взаимосвязи у пеникулин и перитрих, в частности восстановить разделение пеникулин на два отряда: Peniculida и Urocentrida (Puytorac et al., 1987; Striider-Kypke et al., 2000). Анализ нескольких Litostomatea (no SS rRNA) показал, что у изученных представителей этой группы в соответствующем гене последовательность нуклеотидов на 100 единиц короче (за счет helix Е23-5 зоны), чем у остальных инфузорий (Wright, Lynn, 1997b; Wright et al., 1997), что является, по-видимому, характерной чертой представителей класса.
Совмещение филогенетических схем, предложенных на основе морфологических данных (Puytorac et al., 1987; Puytorac et al., 1994; Foissner, 1996; Lynn, Small, 2000; Lynn, 2003) с дендрограммами, построенными в результате сравнения рРНК (Wright, Lynn, 1997; Wright et al., 1997b; Shin et al., 2000; Striider-Kypke et al., 2000), показывает совпадение основных позиций и взаимосвязей групп внутри типа.
Безусловно, для наиболее полного представления об эволюции и филогении Ciliophora, по-видимому, необходимо использовать в сочетании с морфологическими и другие (помимо рРНК) молекулярные признаки (гены): тубулин, гистоны, хитшоковые белки. Число таких работ пока минимально (Bernhard, Schlegel, 1998; Budin, Philippe, 1998).
Систематический обзор
Одноклеточные эукариоты с ядерным дуализмом (макронуклеус и микронуклеус) и ресничным аппаратом, имеющимся на всех стадиях жизненного цикла, а у части сидячих форм — только на стадии мигранта. Соматические кинетосомы одиночные (монокиноиды - моносомы) или парные (дикинетиды = дисомы), обычно собраны в ряды (кинеты = сомакинеты), несимметричные и полярные, совокупность которых (кинетом) характерна для каждого таксона, и у подвижных форм служит основным определительным признаком. С кинетосомами связан сложный комплекс фибрилл. Кинетом делится на 3 зоны: ротовой комплекс (кинеты, мембраны, мембранеллы), соматическая цилиатура (реснички или циррусы поодиночке или в рядах) и тигмотактический аппарат — участки кинет или комплексы кинет и их фрагментов для сигнализации о субстрате или для временного прикрепления к субстрату.Питание разного типа, чаще всего фаготрофное; осмотрофия (точнее, пиноцитоз) встречается у многих групп клеточных или тканевых паразитов — у сукторий рода Endosphaera, у всех Astomatida, у потерявших ротовой аппарат тетрахименид Curimostoma и Dogielella. Характерно наличие трихоцист, трихитов, немадесм и других фибриллярных дериватов кинетосом, образующих опорный и защитный аппараты. Ядра нескольких типов: неделящиеся пара- диплоидные у примитивных форм (отряд Karyorelictida), полиплоидные у высших, гомеомерные и гетеромерные (с полярными или концентрическими зонами «хроматина»). Обновление ядерного аппарата происходит путем автогамии или конъюгации, изогамной или анизогамной, с временным соединением или реже с полным слиянием двух клеток. Дифференцировка вида на изолированные группы клонов по типам спаривания (сингены) доказана у немногих видов гименостомат (тетрахимены), пеникулид (парамеции), гипотрих (эуплотесы).
Размножение делением или почкованием, всегда со сложными процессами морфогенеза; в онтогенезе могут рекапитулироваться предковые признаки, например тетрахимениум у скутицид. Выявлено несколько способов образования ротовой цилиатуры (стома- тогенез), свойственных для разных отрядов цилиат. Почкование характерно для части паразитов (астоматы, гистероцинетиды, ринходиды рода Sphenophrya, апостоматы отряда Pilisuctorida), и для многих сидячих (седентарных) групп — для всех хонотрих и для большинства сукторий; перитрихи диморфные, мигранты могут резко отличаться от трофонтов.
Свободноживущие формы, комменсалы и паразиты. Свободные виды заселяют все типы водных биотопов (моря, реки, озера, Полота, теплые источники, черный ил без кислорода), обычны в почве, во мхах; недавно открыта богатая фауна в воде бромелие- ных растений Южной Америки с большим числом еще не изученных эндемичных родов и видов. Каждый биотоп заселен характерным набором видов. Комменсалы обитают на покровах хозяев или и полостях, широко открытых во внешнюю среду (ротовая, мантийная, жаберная полости); паразиты — это виды, питающиеся тканями и полостными соками хозяина или обитатели пищевого тракта (желудок, кишечник) независимо от их способа питания. Сами цилиаты могут быть комменсалами и паразитами на других цилиатах, к примеру суктории родов Endosphaera, Pseudogemma, Tachyblaston, Riftus, Tokophryella — паразиты сукторий и перитрих.
Цилиаты — древняя группа, судя по распространению на всех континентах и поразительной дивергенции; в ископаемом состоянии известны пелагические морские тинтинниды, как исключение, некоторые фолликулиниды и перитрихи-лорикаты; несколько без- раковинных мобильных видов, например Paramecium triassicum, найдено в янтаре. Количество видов в подтипе определяется цифрой 8000, но только одна недавняя экспедиция в южноафриканскую полупустыню Намиб обнаружила несколько сотен новых видов и 37 новых родов. Долгое время господствовало мнение о космополитном распространении всех не-симбионтных цилиат, но это неверно, есть множество локальных эндемиков и видов, встречающихся только на южных континентах.
Изученность симбионтов беспозвоночных ничтожна. Следует ожидать значительного роста числа видов и родов симбионтных и свободных цилиат в ближайшей перспективе. В систематике цилиат характерна тенденция к дроблению родов, выделяемых у гапто- рид, никтотерид, гипотрих и гименостомат и в других группах, нередко по ничтожным признакам и поэтому трудно определимых; подроды за редкими исключениями у цилиат не выделяются.
Книги:
 Протисты. Руководство по зоологии. Часть 1. Алимов А.Ф. 2000
Протисты. Руководство по зоологии. Часть 1. Алимов А.Ф. 2000
 Протисты. Руководство по зоологии. Часть 2. Алимов А.Ф. 2007
Протисты. Руководство по зоологии. Часть 2. Алимов А.Ф. 2007
 Протисты. Руководство по зоологии. Часть 3. Алимов А.Ф. 2011
Протисты. Руководство по зоологии. Часть 3. Алимов А.Ф. 2011
 Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987
Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987



 Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
 Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423
Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423
 Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989
Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989