



ОПИСАНИЕ:
Отряд Арцеллообразные / Arcellinida Kent, 1880
Корненожки, имеющие раковины или иные покровные структуры с 1 устьем, построенные из органического или органического и неорганического материала. Пресноводные.Морфология и физиология
СТРОЕНИЕ КЛЕТКИ
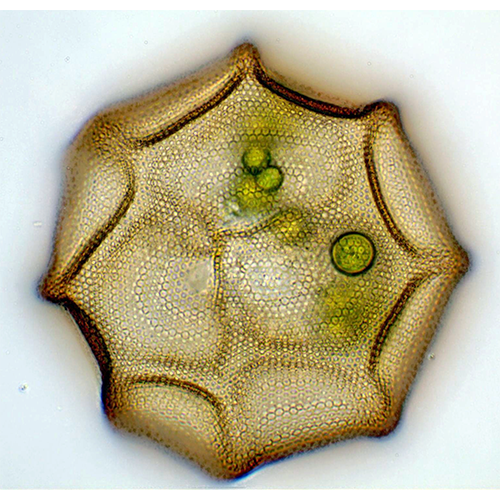
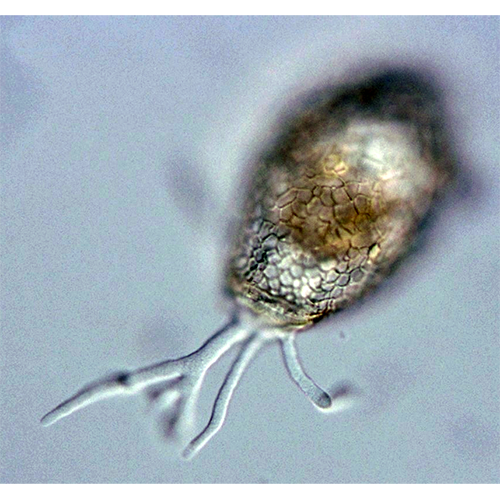
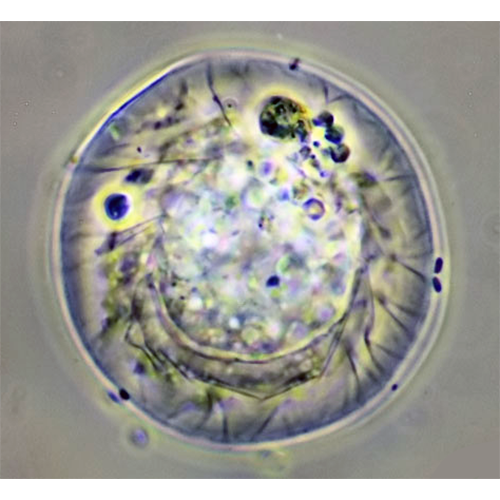
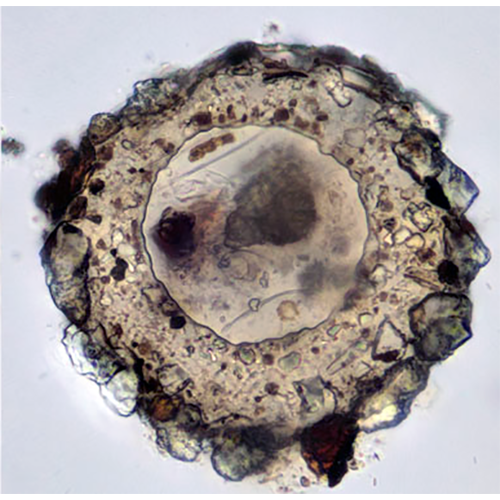
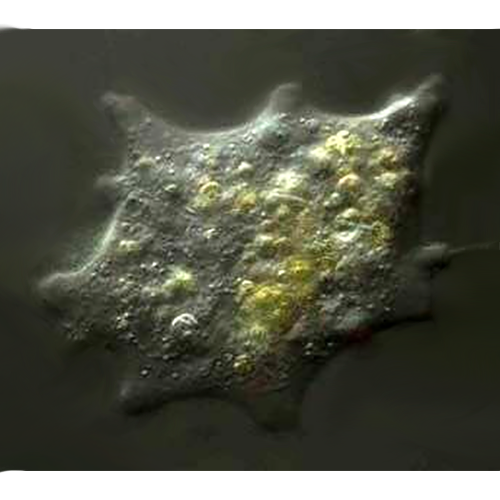
У раковинных амеб собственно амебоидная клетка заключена в раковину («домик»), которая состоит из агглютинированного клеткой экзогенного материала или из секретированных ею частиц. Тело амебы, как правило, не заполняет полость раковины целиком, а закреплено в ней при помощи эпиподий — выростов цитоплазмы, прикрепленных изнутри к стенке раковины. Иногда цитоплазматическое тело амебы соединяется с внутренней поверхностью раковины лишь в области устья, по его периферии.Снаружи клетка покрыта плазмалеммой, в состав которой входит тонкий слой недифференцированного гликокаликса. В цитоплазме находится ядро (ядра) и набор органелл, типичный для аэробных корненожек.
Ядра раковинных амеб обычно везикулярного типа, с более или менее компактным ядрышком, реже — гранулярного типа. Большинство представителей отряда одноядерные, но для некоторых характерно наличие 2 ядер; известны также многоядерные виды (Arcella megastoma). У изученных представителей родов Arcella, Hyalosphenia, Nebela, Lesquereusia оболочка ядра изнутри армирована слоем фибриллярного материала. Деление ядра на ультраструктурном уровне изучено только у Arcella vulgaris, у которой оно носит характер закрытого ортомитоза.
Митохондрии имеют тубулярные кристы, которые могут анастомозировать. Матрикс митохондрий может быть электронно-плотным (у изученных представителей родов Cryptodifflugia, Hyalosphenia, Centropyxis) или электронно-светлым (Lesquereusia, Netzelia). У разных представителей одного и того же рода митохондрии могут иметь или плотный, или светлый матрикс, однако гетероморфизм митохондрий у раковинных амеб неизвестен.
Для Testacealobosia характерно наличие в цитоплазме многочисленных диктиосом аппарата Гольджи. Он принимает участие в синтезе цементных везикул, содержимое которых скрепляет частицы, входящие в состав стенки раковины. У видов, имеющих секретированную раковину, аппарат Гольджи участвует в синтезе идиосом для ее построения.
В клетке раковинных амеб обычно имеется 1 или 2 сократительные вакуоли; известны также виды, у которых их число достигает нескольких десятков. Вакуоли опорожняются либо в полость раковины, либо наружу.
Шероховатая эндоплазматическая сеть (ШЭПС) у многих видов образует плотные скопления в цитоплазме, которые находятся около ядра, а иногда — окружают его. Эти скопления эндоплазматической сети прокрашиваются гематоксилином и другими красителями, окрашивающими рибонуклеопротеиды, и иногда совершенно маскируют ядро. В литературе начала века их часто описывали под названием «хромидии» и рассматривали даже как одну из альтернативных форм существования ядра клетки у раковинных амеб.
У видов, которые синтезируют для построения стенки раковины кремниевые чешуйки, известна модификация ШЭПС — силикалемма. В ее цистернах происходит синтез идиосом. Некоторые раковинные амебы способны покрывать тонким слоем кремния фагоцитированные частицы, например бактерии. Впоследствии эти частицы встраиваются в стенку раковины.
В составе цитоскелета раковинных амеб выявлены микрофиламенты и, по крайней мере у некоторых видов, микротрубочки. Пучки микрофиламентов обнаружены в псевдоподиях движущихся амеб; их скопления имеются в различных участках цитоплазмы. Микротрубочки в псевдоподиях не отмечены, однако у некоторых видов рода Arcella они ассоциированы с поровыми комплексами и располагаются вокруг ядра.
В цитоплазме клеток находится большое количество различных включений, которые представляют собой кристаллы (в основном оксалаты), липидные глобулы, цементные везикулы и материал для построения дочерней раковины. В цитоплазме большинства изученных видов наряду с митохондриями находятся микротельца, окруженные одинарной мембраной.
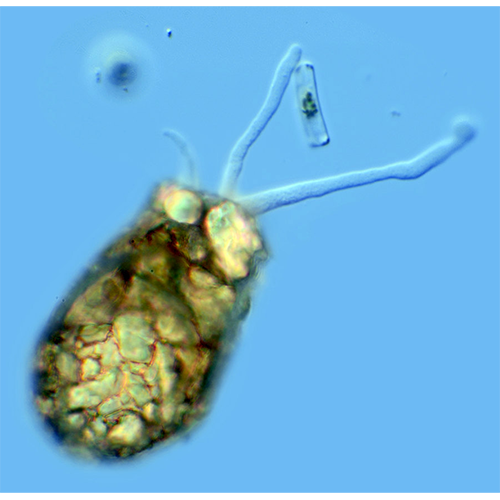
Многие раковинные амебы факультативно или облигатно содержат в цитоплазме различные эндоцитобиотические организмы (например, Difflugia gramen, D. lobostoma, Hyalosphenia papilio, Cucurbitella mespiliformis). Как правило, это зоохлореллы — одноклеточные зеленые водоросли из рода Chlorella. Иногда в цитоплазме амеб отмечали наличие водорослей из родов Protococcus и Scenedesmus. Симбиоз с хлореллами у некоторых видов носит, видимо, облигатный характер, в то время как другие могут обходиться и без симбионтов. Показано, что виды, содержащие хлореллы, гораздо дольше других раковинных амеб могут обходиться без пищи (некоторые крупные виды диффлюгий — до 3 мес). При помещении их в темноту водоросли постепенно исчезают из цитоплазмы и амебы вскоре гибнут. Бактерии в цитоплазме пока обнаружены только у Difflugia geosphaera. Они лежат свободно в цитоплазме клетки, однако характер взаимоотношений амеб с бактериями неясен.
Наличие раковины, придающей более или менее стабильную форму цитоплазматическому телу клетки, создает предпосылки для компартментализации цитоплазмы — разделения ее на различные в функциональном отношении участки. Это разделение хорошо выражено у Testacealobosia, имеющих округлую или грушевидную раковину (представители рода Difflugia и морфологически схожих с ним родов), и практически отсутствует у амеб с уплощенной раковиной (типа раковин Arcella spp. и Centropyxis spp.).
У амеб с вытянутой или округлой раковиной ядро всегда располагается в задней (аборальной) ее части. Вокруг него обычно находится плотное скопление ШЭПС. Этот участок цитоплазмы иногда называют «зоной плотной ЭПС». На периферии этой части клетки располагаются диктиосомы, иногда — запасные идиосомы и сократительные вакуоли. Никаких других органелл и пищеварительных вакуолей здесь нет.
Ближе к устью (в оральной части цитоплазмы) располагаются все митохондрии, вакуоли, кристаллы, цементные везикулы и частицы, фагоцитированные клеткой для построения дочерней раковины. Псевдоподии начинают формироваться несколько глубже границы устья раковины, и поэтому ближайшая к нему часть цитоплазмы представляет собой сеть переплетающихся и анастомозирующих тяжей.
РАКОВИНА.
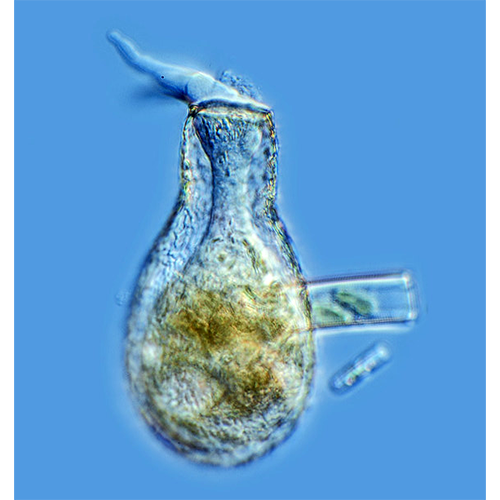
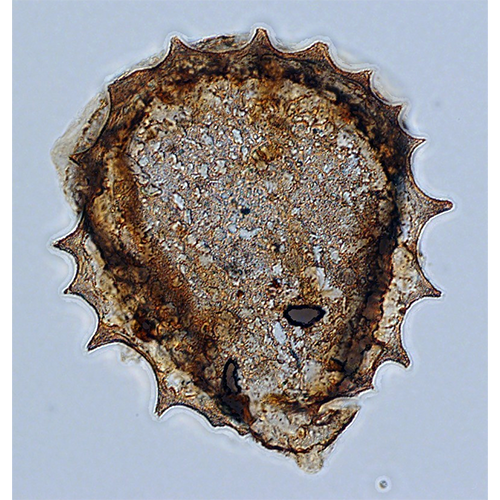
Подавляющее большинство Testacealobosia имеют жесткую раковину, которая не способна к какой- либо конформации. Исключением является представитель рода Microchlamys — М. patella, раковина которого представляет собой находящийся на спинной стороне тела амебы мягкий щиток, который может «складываться». Этот щиток образован одним слоем гексагональных субъединиц — альвеол. Цитоплазматическое тело этой амебы покрыто тонким мембранным мешком, который прикреплен к внутренней поверхности щитка и имеет центральное отверстие для выхода псевдоподий. Такая организация является исключением, у всех остальных раковинных амеб подобного мешка нет.Форма раковин чрезвычайно разнообразна — они могут быть дисковидными, колпачковидными, овальными, грушевидными, полусферическими или почти сферическими. Края латерально сжатых или уплощенных раковин бывают окружены килем разной ширины.
Если раковина уплощена, то часть ее, на которой находится устье, называется брюшной поверхностью, а противоположная — спинной. У раковин, удлиненных по оси, проходящей через центр устья, различаются передняя (устьевая) и задняя части. Подобное же разделение раковин на приустьевую, переднюю и заднюю части применяется и в отношении раковинных амеб, имеющих уплощенную брюшную поверхность и эксцентрично расположенное устье. При этом если задняя часть раковины вздута, то она называется брюшком, если же небольшое вздутие имеется и в передней части раковины, то для его обозначения употребляется термин «козырек».
Раковины Arcellinida однокамерные, хотя внутри них могут встречаться структуры, подразделяющие их полость. Внутри раковины Pontigulasia проходит узкий мостик, построенный из органического материала. У близких родов Zivcovicia и Lagenodifflugia уже имеется плотная диафрагма, являющаяся продолжением внутренней стенки раковины и отделяющая полость основной части раковины от шейки; для выхода псевдоподий в этом случае служат 1 или 2 отверстия в диафрагме. Диафрагма имеется и у Lesque- reusia spiralis. У многих почвенных раковинных амеб (например, у Plagiopyxis) вентральная часть раковины глубоко вдается в ее полость, отделяя брюшко от ограниченной козырьком камеры; в этом случае устье лежит в глубине раковины.
Форма и расположение устья у раковинных амеб чрезвычайно широко варьируют. Устье может быть круглым, эллипсоидным, трех-, четырех- й многолопастным, зубчатым, щелевидным, иметь форму восьмерки и т. д. Устье может располагаться в нижнем конце вытянутой раковины (Difflugia), в центре брюшной поверхности (Arcella, Cyclopyxis) или эксцентрично (Centropyxis. Plagiopyxis). Оно может находиться непосредственно на поверхности раковины (Phryganella), либо на конце так называемой устьевой воронки — впячивания вентральной поверхности, вдающегося внутрь раковины (Cyclopyxis, Centropyxis, Arcella). У некоторых видов рода Arcella устье располагается на конце устьевой трубки, направленной вниз от конца устьевой воронки. Иногда устье открывается в камеру, ограниченную козырьком. Встречаются раковинные амебы с крупным устьем (его диаметр может составлять больше половины диаметра раковины) так называемые эвристомные формы, и микростомные раковинные амебы, у которых округлое или радиально-симметричное устье оказывается значительно суженным. Уменьшение эксцентрично расположенного устья приводит к формированию щелевидного устьевого отверстия. У представителей сем. Mississippiellidae устье образовано серией из 10—18 мелких овальных отверстий, располагающихся дуговидными рядами. Устье может быть окружено валиками, воротничками, края устья могут утолщаться или заходить один на другой, образуя «губы»; приустьевая часть может вытягиваться в удлиненную шейку.
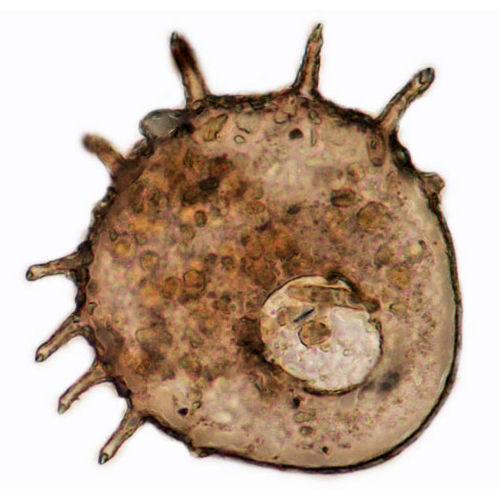
На раковинах Arcellinida могут встречаться многочисленные выросты. Различают шипы (выросты, в которые заходит полость раковины) и не имеющие внутренней полости иглы.
По общим чертам строения можно выделить следующие морфологические типы раковин раковинных амеб.
1. Акростомный — устье терминальное, ось симметрии раковины проходит через центр устья. В этом типе различаются простые акростомные (радиально-симметричные), уплощенные акростомные и изогнутые акростомные раковины, характеризующиеся билатеральной симметрией.
2. Трахелостомный — с устьем на шейке, либо располагающейся на продольной оси раковины, либо изгибающейся и отделенной сужением от расширенного брюшка.
3.Аксиальный — раковина в виде части сферы с уплощенной вентральной поверхностью, центр которой Занят устьем. Устье может быть округлым, треугольным, зазубренным. Иногда имеется устьевая воронка, вдающаяся в глубь раковины.
4. Эллипсостомный — эллипсоидное устье располагается в центре уплощенной вентральной поверхности раковины, имеющей овоидные очертания. Ось устья совпадает с продольной осью вентральной поверхности раковины.
5. Пропилостомный — устьевая воронка как бы образует переднюю камеру, открывающуюся снаружи на вентральной поверхности раковины, а внутри заканчивающуюся собственно устьем.
6. Диплостомный — имеется обширная вестибулярная полость, ограниченная со стороны вентральной подошвы диафрагмой с 2 почковидными отверстиями. Собственно устье располагается в верхней части вестибулярной полости. Имеется скелетная система, укрепляющая эту полость.
7. Плагиостомный — с эксцентричным устьем, лежащим на вентральной подошве, что определяет билатеральную симметрию раковины. Возможно развитие устьевой воронки, иногда с «уздечками», соединяющими ее с дорсальной стенкой раковины.
8. Плагиостомный с козырьком — устье открывается в камеру, ограниченную козырьком. Отверстие этой камеры располагается на подошве раковины.
9. Криптостомный — сходный с плагиостомным, но устье имеет резко выраженную тенденцию к уменьшению размеров и обычно принимает вид узкой поперечной щели.
10. Криптостомный с козырьком — отличается от плагиостомного с козырьком крайней редукцией дистального отверстия ограниченной козырьком камеры до линейной или полулунной щели.
У некоторых видов Arcella устье окружено порами. Поры могут находиться и в апикальной части раковины (Hyalosphenia и Nebela). Их функциональное значение остается неясным. Во время эксцистирования у Nebela tincta наблюдали выход псевдоподий через латеральные поры.
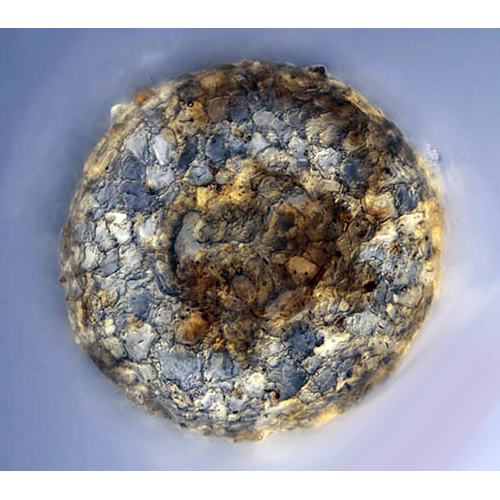
По способу построения раковины подразделяются на агглютинированные и состоящие из гомогенного материала. Стенка агглютинированных раковин состоит из частиц, скрепленных органическим цементом (матриксом), который синтезируется клеткой. Частицы могут быть фагоцитированы амебой из окружающей среды (ксеносомы) или секретированы самой клеткой (идиосомы и протеинозные альвеолы). Иногда в состав стенки раковины входят как ксеносомы, так и частицы, синтезированные клеткой. Раковины, состоящие из гомогенного материала, обычно образованы несколькими органическими или минеральными слоями.
Амебы, строящие агглютинированные раковины, зависят от имеющегося в окружающей среде строительногоматериала и поэтому в разных условиях могут иметь различную их структуру. Как правило, в качестве ксеносом используются песчинки и панцири диатомовых водорослей. Преобладающими минеральными компонентами таких раковин являются кремнезем и кварц. Многие виды Testacealobosia, видимо, обладают способностью к выбору определенных частиц для построения раковины: одни виды рода Difflugia строят гладкие раковины из небольших уплощенных кварцевых частиц, в то время как другие формируют шероховатые раковины из относительно крупных частиц. Некоторые виды в качестве материала для построения раковины используют преимущественно панцири диатомовых водорослей.
У ряда раковинных амеб (например, у Heleopera) преобладающим компонентом раковины являются чешуйки фагоцитированных ими мелких филозных раковинных амеб, в основном из рода Euglypha. При культивировании в условиях, исключающих возможность питания этими амебами, Heleopera spp. строит раковину только из органического материала. У некоторых раковинных амеб (например, Centropyxis spp.) встречаются все переходы от органических раковин к агглютинированным. Экзогенными частицами у них часто покрыта только спинная сторона раковины.
Эндогенные минеральные частицы (идиосомы) представляют собой кремнеземные или известковые образования различной формы. У Nebela они имеют вид чешуек различной формы. У Lesquereusia spiralis они палочковидные, слегка изогнутые, а у Netzelia представлены небольшими округлыми частицами Paraquadrula идиосомы состоят из извести и имеют вид пластинок, а у Mississippiella они образованы ромбоидными кристаллами кальцита.
Раковину из протеинозных альвеол, скрепленных матриксом, имеют Arcella, Centropyxis, Microchlamys и некоторые другие роды. Альвеолы — это полые пентагональные образования. В качестве основного компонента в их состав входит вещество белковой природы, близкое к кератину.
Показано, что раковина Arcella содержит аминокислоты: глицин, аланин, лейцин, аргинин, валин, фенилаланин, цистеин, тирозин и глутаминовую кислоту. В старых раковинах альвеолы обычно заполнены неорганическими соединениями, например солями железа. Наружная поверхность может быть покрыта дополнительным слоем матрикса.
ПСЕВДОПОДИИ И ДВИЖЕНИЕ.
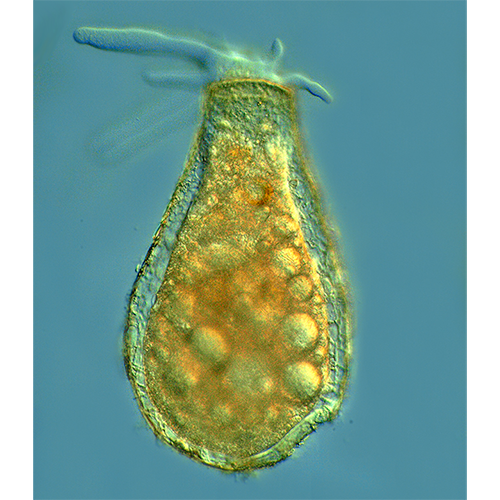
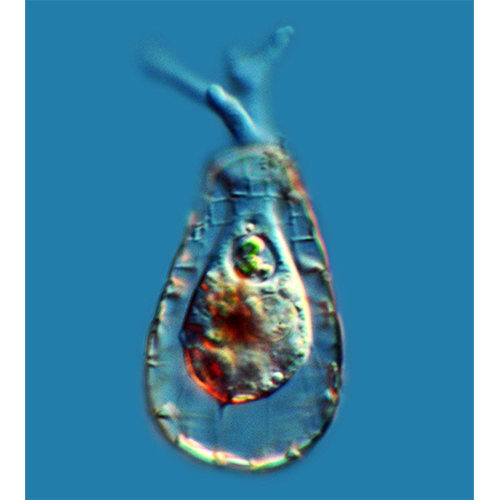
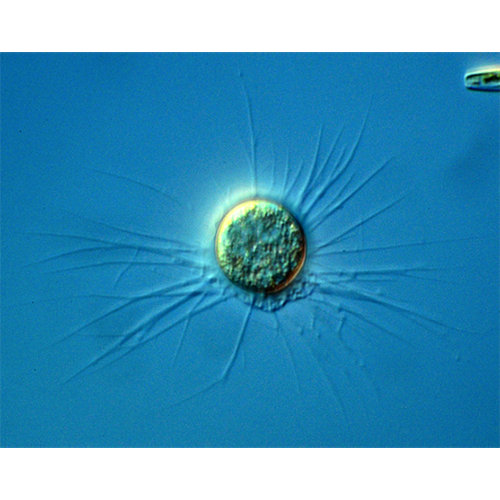
Перемещение и захват различных частиц осуществляются при помощи псевдоподий. Иногда выделяют 2 их типа: экзолобоподии, состоящие только из гиалоплазмы (особой их формой являются ретикуло-( лобоподии — заостренные на концах и разветвляющиеся с образованием анастомозов), и эндолобоподии, в состав которых входит и гиало- и гранулоплазма.Мелкие и средние раковинные амебы при передвижении обычно поднимают домик вертикально. Цитоплазма при таком движении растекается по субстрату, образуя вентральный валик, на котором ползет амеба. Подобное перемещение очень характерно для Centropyxis, Arcella, мелких диффлюгий.
Амебы, имеющие крупные агглютинированные домики, очень редко отрывают их от субстрата. Как правило, при движении такая амеба образует длинные псевдоподии, концы которых прикрепляются к субстрату. Потом псевдоподии втягиваются, подтаскивая раковину.
Скорость движения раковинных амеб различна, но в целом это малоподвижные организмы. Скорость перемещения Arcella vulgaris 51—90 мкм/мин, т. е. 3— 5.5 мм/ч; Difflugia crehulata перемещается со скоростью 60 мкм/мйн; Quadrulella symmetrica — 32 мкм/мин.
Питание и осморегуляция
Большинство раковинных амеб — полифаги. Они питаются бактериями, водорослями, спорами грибов, жгутиконосцами, инфузориями или даже коловратками. В пищеварительных вакуолях почвенных раковинных амеб часто обнаруживаются частицы отмерших растений. Неоднократно описывалась способность раковинных амеб к пробуравливанию стенок водорослей, вслед за чем в образовавшееся отверстие клетка просовывает псевдоподии и водоросль «выедается» изнутри. Некоторые виды (Arcella arenaria, A, polypora) — облигатные фитофаги.Отмечены случаи питания некоторых раковинных амеб другими раковинными амебами. Так, например, Heleopera и Nebela фагоцитируют Euglypha и Trineта (класс Filosea) и в дальнейшем встраивают идиосомы заглоченных раковин в собственную раковину. Мелкая Hyalosphenia platystoma прикрепляется к раковинам крупных раковинных амеб, проделывает сбоку отверстие и опустошает их.
Интенсивность питания достаточно высока — интервал между заглатыванием отдельных пищевых частиц у некоторых видов составляет 4—7 мин. Продолжительность переваривания пищи составляет 30—60 мин, фаза дефекации длится 2—3 мин. При дефекации псевдоподии втягиваются, вакуоль с непереваренными остатками подходит к мембране в области устья й выбрасывает свое содержимое во внешнюю среду.
Сократительные вакуолй у раковинных амеб работают с разной интенсивностью в зависимости от условий окружающей среды, но обычно интервал между систолами составляет 20—60 с.
Размножение и жизненные циклы
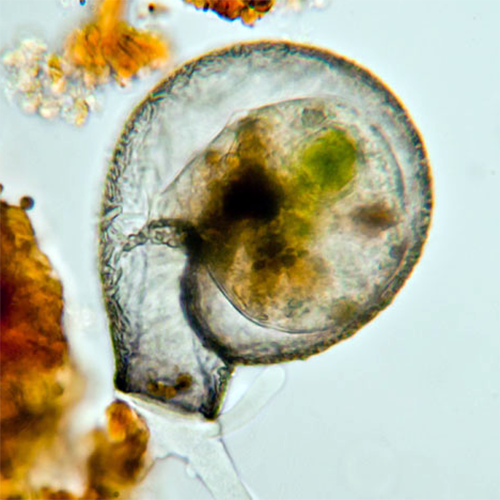
Размножение происходит путем бинарного деления. Детали этого процесса отличаются у представителей различных родов, однако в целом можно выделить 2 наиболее характерных типа морфогенеза дочерней особи.У Arcella и других раковинных амеб, имеющих органическую альвеолярную раковину, при подготовке к делению в цитоплазме накапливаются так называемые текагенные гранулы — везикулы, содержащие в упакованном виде альвеолы для построения дочерней раковины. У Arcella диаметр этих гранул около 500 нм, а высота до 900 нм; их общий объем может составлять до 7 % объема цитоплазмы клетки.
У Arcella vulgaris процесс деления начинается с того, что часть цитоплазмы выпячивается из устья материнской раковины, образуя текагенную почку. По размеру она примерно равна материнской особи. Текагенные гранулы перемещаются к поверхности этой почки и располагаются под ней в 1 слой. На этом этапе клетка секретирует на поверхность почки слой гомогенного вещества. Вслед за этим происходит синхронный выброс содержимого гранул. Оказавшись на поверхности, альвеолы увеличиваются в размерах и сливаются друг с другом. Видимо, первоначальное скрепление альвеол происходит за счет ранее секретированного вещества.
Затем амеба образует куполообразную псевдоподию, которая окружает только что образовавшуюся раковину. Внутреннее пространство этого купола определяет окончательную форму й размеры дочерней раковины. Увеличение размеров дочерней раковины происходит за счет движений цитоплазмы текагенной почки, которая «расталкивает» ее изнутри. Затем цитоплазматический купол быстро втягивается. Весь процесс формирования дочерней раковины занимает около 10 мин.
В результате взаимного сжатия альвеол формируется гексагональная структура стенки дочерней раковины. В местах, где соприкасаются 3 альвеолы, образуются микроскопические поры. Системой таких пор оказывается окружена каждая альвеола.
На последних этапах деления дочерняя и материнская раковины соединены друг с другом специальным замком из содержимого текагенных гранул. Он имеет вид муфты, проходящей через устья раковин и «зацепленной» за края устий изнутри. Когда завершается распределение цитоплазмы между клетками, замок разрушается и происходит окончательное деление клеток.
В цитоплазму дочерней клетки мигрируют митохондрии и другие органеллы. Происходит митотическое деление ядра, и одно из ядер перемещается в дочернюю клетку. Разрушается замок, происходит окончательное разделение цитоплазмы, и дочерняя клетка переходит к самостоятельному существованию. После достройки раковины цитоплазма отделяется от ее стенок и образуются эпиподии, на которых подвешено тело взрослой особи.
Подобный ход морфогенеза характерен для всех раковинных амеб, строящих раковину из органических альвеол. Однако формирование цитоплазматического купола при морфогенезе подробно прослежено только у Arcella.
У молодых раковин каждая альвеола имеет внутреннюю полость, с возрастом просвет альвеол часто заполняется неорганическим веществом: у арцелл это железо; у Centropyxis hirsuta —марганец. С накоплением неорганических веществ связана более темная окраска старых раковин.
У раковинных амеб, строящих агглютинированные раковины, делению предшествует период активного поглощения ксеносом. В начале деления из устья материнской раковины выпячивается участок цитоплазмы. Он быстро увеличивается в размерах и принимает форму, примерно соответствующую форме будущей раковины. К поверхности этого выпячивания перемещаются вакуоли с ксеносомами и с органическим цементом. Иногда амеба образует пальцевидные, весьма подвижные псевдоподии, которые могут захватывать частицы из внешней среды. Происходит экзоцитоз содержимого вакуолей, и ксеносомы скрепляются органическим цементом. Крупные частицы из внешней среды могут просто «прилипать» к стенке дочерней раковины. Ядро претерпевает деление, часть органелл и одно из ядер мигрируют в дочернюю клетку. Вслед за этим происходит окончательное разделение цитоплазмы клеток. Дочерняя клетка еще некоторое время достраивает раковину, фагоцитируя ксеносомы и включая их в стенку раковины. Изнутри раковина выстилается слоем органического цемента; за счет него также формируется органическая каемка вокруг устья и заполняются промежутки между частицами. У взрослой особи цитоплазма.отделяется от стенок раковины и образуются эпиподии.
У амеб, которые включают в состав раковины наряду с ксеносомами еще и идиосомы, морфогенез раковины проходит по той же схеме, хотя его детали могут варьировать у разных видов.
В литературе имеются описания процесса, названного экзувиацйей. У Hyalosphenia papilio иногда из устья раковины выпячивается цитоплазма. Объем этого выроста растет, в то время как объем цитоплазмы в раковине уменьшается. На поверхности этого выпячивания формируется раковина, после чего в нее перетекает вся цитоплазма клетки. Ядро переходит в новую раковину без каких-либо изменений; деления не происходит. Старая раковина остается пустой. Некоторые современные исследователи подвергают сомнению существование процесса экзувиации.
Часто можно встретить описание конгрегации — объединения 2 раковинных амеб, слияния их цитоплазматических тел и последующего инцистирования продукта слияния. Однако ни одному исследователю не удалось пока достоверно наблюдать эксцистирование после подобного процесса.
Покоящиеся стадии раковинные амебы образуют при наступлении неблагоприятных условий (изменение температуры, понижение влажности). При этом псевдоподии втягиваются, сократительная вакуоль прекращает свою деятельность. Устье раковины закрывается эпифрагмой, представляющей собой прозрачную органическую мембрану, или пробкой из ксеносом или идиосом. Внутри такой раковины часто находится «предцистная» форма амебы (псевдоциста), которая отличается отсутствием цистной оболочки. Истинная циста покрыта плотной защитной оболочкой, в состав которой могут входить и ксеносомы. Как правило, циста сферическая или слегка уплощенная. Эксцистирование происходит при попадании цисты в благоприятные условия.
Arcella vulgaris помимо обычных, покоящихся цист образует особые мейотические цисты, в которых происходит половой процесс, протекающий по типу автогамии. Мейотическое деление ядра при этом прослежено на ультраструктурном уровне. Из 8 образующихся ядер 4 пикнотизируются и перевариваются, а оставшиеся сливаются попарно, восстанавливая исходное двухъядерное состояние клетки. Достоверных наблюдений полового процесса у других раковинных амеб пока нет.
Жизненный цикл Testacealobosia, как правило, представляет собой серии бинарных делений, которые прерываются инцистированием, происходящим при наступлении неблагоприятных условий. У Arcella vulgaris в него включается половой процесс, однако происходит он регулярно или нет, пока неясно. По-видимому, у некоторых видов в него могут нерегулярно включаться конгрегация и экзувиация, однако все эти процессы нуждаются в более подробном описании.
Для одного из видов раковинных амеб — Difflugia limnetica, характеризующегося чередованием в жизненном цикле бентической и планктонной фаз, было описано явление цикломорфоза. Во время бентического периода жизни раковина состоит из мелких песчинок и воротничок у нее развит слабо, а в эпилимнионе в состав раковины входят панцири диатомей и имеется развитый воротничок.
Экология и распространение.
УСЛОВИЯ ОБИТАНИЯ.
Отряд Arcellinida составляют преимущественно пресноводные организмы, лишь небольшая их часть обитает в интерстициали морских побережий. Можно утверждать, что раковинные амебы встречаются почти всюду, где имеется доступная для них влага, — от крупных озер и рек до пространств между частицами почвы.В крупных пресноводных водоемах можно говорить об отдельных сообществах раковинных амеб, характерных для фитали и профундали. В фитали преобладают раковинные амебы с мелкими раковинами (Paraquadrula, Cryptodifflugia, Phryganella, мелкие виды рода Difflugia), формы с раковинами из органического материала, с идиосомами или с редкими мелкими инкрустирующими частицами. В этой зоне часто встречаются формы с уплощенными в дорсовентральном направлении раковинами; относительно большой процент видов имеет шипы или иглы.
С увеличением глубины и с переходом к обитанию на поверхности донных осадков увеличивается количество видов с крупными раковинами, появляется большое число форм с крупными инкрустирующими частицами, преобладают шаровидные или округлые в сечении раковины без шипов и игл. В зоне, лишенной растительности, население раковинных амеб донных осадков слабо зависит от глубины водоема. В профундали обычно доминирует род Difflugia, наряду с которым могут встречаться представители родов Pontigulasia, Lesquereusia, Heleopera, Phryganella.
В настоящее время доказано, что существует 3 основных сообщества пресноводных раковинных амеб, которые позволяют судить о характере водоема и о типе его осадков. Эти сообщества характерны для типичных олиготрофных озер с богатыми кислородом придонными слоями, эвторфных озер с отложениями гиттии и недостатком кислорода в придонных слоях и для небольших озер с низким значением pH воды и с отложениями дью на дне. Кроме этих 3 встречаются и смешанные сообщества. Следует отметить, что большая часть пресноводных раковинных амеб представлена так называемыми индифферентными видами, не характерными для какого-либо определенного типа водоемов.
В болотах раковинные амебы бывают связаны преимущественно со сфагнумом, причем заселяют в основном пазухи листочков мха. Имеются торфобионтные виды, часто встречающиеся в болотах, и торфоксенные виды, обнаруживаемые там лишь изредка. К типичным торфобионтным видам относятся Difflugia bacillifera, Hyalosphenia papilio, H. elegans, Nebela tenella. Для группы болотных раковинных амеб также показано наличие комплексов видов, присущих определенным типам болот. Широко известна классификация сообществ раковинных амеб, предложенная Харнишем (Harnisch, 1927), который выделяет 5 таких комплексов, встречающихся в различных типах или участках болот. Основным фактором, определяющим характер этих сообществ, является влажность среды. В пределах одного болота раковинные амебы обычно распределяются в прямой зависимости от влажности мха, причем по мере уменьшения влажности увеличивается количество микростомных форм.
Еще более четко выражена зависимость от влажности среды сообществ раковинных амеб, обитающих в лиственных мхах. И в этом случае можно проследить связь микростомии раковинных амеб с уменьшением влажности окружающей среды.
Исследование вертикального распределения раковинных амеб в пределах моховой подушки показывает, что можно выделить 3 зоны: верхний, зеленый слой мха, практически не заселяемый раковинными амебами; слой с отмирающими листочками, в котором встречается большое количество преимущественно уплощенных плагиостомных видов, среди которых редки формы, строящие раковины из ксеносом; слой почвы, пронизанной ризоидами мха, в котором преобладают крупные сфероидные формы с раковинами, состоящими из большого количества ксеносом.
В почве обитает большое количество видов раковинных амеб, образующих там иногда довольно плотные популяции. Наиболее густо бывает заселена подстилка, в которой благодаря наличию системы чрезвычайно тесных полостей (между отдельными листьями в опаде) преобладают раковинные амебы с уплощенными раковинами. Для фауны раковинных амеб подстилки характерно преобладание видов с органическими раковинами либо с раковинами, построенными из идиосом. При переходе к гумусовому горизонту полости между отдельными почвенными частицами становятся значительно больше, вследствие чего в этом слое преобладают раковинные амебы с относительно крупными, объемистыми, часто сферическими или близкими к этому типу раковинами. В фауне данного горизонта преобладают раковинные амебы с раковинами, построенными Из ксеносом, причем здесь встречается большое количество специфических терробионтных видов, для которых характерна отчетливо выраженная тенденция к крип- тостомии. Только для почв характерны многие виды рода Plagiopyxis, Centropyxis cryptosoma, С, vandeli и другие виды.
В интерстициальной фауне отмечаются некоторые эврибионтные представители отряда Arcellinida. Единственный характерный только для интерстициа- ли род отряда — Pomoriella (сем. Hyalospheniidae). Очень часто в пробах песка, взятых на морских побережьях, встречается Centropyxis constricta, рассматриваемый на этом основании как псаммофильный вид.
Малые размеры раковин амеб и их способность к цистообразованию обеспечивают возможность их легкого распространения. Распространение раковинных амеб при помощи ветра было доказано их отсутствием в свежевыпавшем снегу и постоянным присутствием пустых раковин или цист в снегу, лежавшем длительное время в горах и на ледниках. Возможно распространение раковинных амеб животными. Ин- цистированные раковинные амебы неоднократно обнаруживались в загрязнениях перьевого покрова птиц и шерстного покрова млекопитающих. Отмечены случаи переноса раковинных амеб панцирными клещами.
ГЕОГРАФИЧЕСКОЕ РАСПРОСТРАНЕНИЕ.
Вопрос о географическом распространении раковинных амеб не является окончательно решенным в первую очередь из-за неравномерного изучения их фауны в разных регионах земного шара. Все основные данные в этой области были получены до признания самостоятельности групп раковинных амеб с лобоподиями и с филоподиями, и поэтому отмеченные закономерности в равной степени относятся как к подклассу Testacealobosia (класс Lobosea), так и к подклассу Testaceafilosia (класс Filosea).Из 1630 видов и подвидов раковинных амеб обоих подклассов, описанных до 1975 г., 1032 были обнаружены в Европе, 684 в Африке, 428 в Южной Америке, 424 в Северной и Центральной Америке, 229 в Азии, 89 в Антарктиде.
Неоднократно выдвигавшаяся гипотеза об абсолютно космополитном распространении раковинных амеб в последние десятилетия подвергается сомнению. Можно утверждать, что не все раковинные амебы являются космополитами, но тенденция к космополитизму выражена у этой группы значительно более резко, чем у многих других групп животных.
Среди раковинных амеб встречаются убиквисты с космополитным распространением. Чрезвычайно широко распространены космополиты, связанные с определенными типами среды, так что в большинстве случаев видовой состав раковинных амеб скорее определяется типом биотопа, чем географическим положением территории. Отмечены виды с рассеянным распространением; есть основания полагать, что существуют отдельные виды, ограниченные в своем распространении определенными географическими зонами. Неоднократно делались попытки выявления комплексов видов, характерных для отдельных географических зон, однако результаты таких работ не могут считаться убедительными, до тех пор пока все эти зоны не будут исследованы хотя бы с примерно одинаковой степенью тщательности.
Была выдвинута гипотеза, объясняющая закономерности распространения некоторых раковинных амеб с позиций теории континентального дрейфа. Ван Ойе (Van Оуе, 1956) удалось выделить комплекс видов, характерный для участков суши, составлявших Гондвану (Южная Америка, Новая Зеландия, Австралия, Антарктида). На примере рода Nebela он показал наличие общих видов в Новой Зеландии и Южной Америке и различие видового состава этого рода в фауне Новой Зеландии и Палеарктики. Проведенный недавно анализ распространения бриофильных видов также позволил выделить 2 крупных комплекса видов, один из которых по своему распространению соответствует границам Гондваны, а другой встречается в Северной Америке, Европе и Азии, что соответствует Лавразии.
Если рассматривать данные по распространению отдельных экологических группировок раковинных амеб, то можно найти определенные закономерности. Считается, что фауна водных раковинных амеб состоит в основном из широко распространенных видов, хотя некоторые авторы допускают наличие местных фаун. Неоднократно подчеркивалась оригинальность тропической фауны. Фауна раковинных амеб мхов и примитивных почв считается космополитной. Исключения составляют Nebela cartesi, N cockayini и Certesella martiali, отмеченные только в южном полушарии, и Nebela vas, встречающаяся в южном полушарии и приэкваториальных зонах северного полушария. В почвах имеются, как правило, 2 фракции: субвоздушная космополитная, близкая к сообществу раковинных амеб мхов, и космополитная эдафическая. К этому комплексу в некоторых случаях добавляются почвенные виды с ограниченным распространением: либо почвенный тропический комплекс, характерный для тропической зоны Африки, Южной Америки, Мексики, Непала, либо почвенный комплекс умеренных и холодно-умеренных зон Канады, высокогорий Мексики и Японии. Фауна почвенных амеб лесного гумуса Европы состоит только из широко распространенных видов.
Раковинные амебы часто встречаются в очень больших количествах и в некоторых биотопах могут являться доминирующей в количественном отношении группой. Максимальная их численность в зоне фиталй олиготрофных озер может достигать 6, в почве — 800 млн/м2. Биомасса раковинных амеб в почве может, по некоторым данным, доходить до 800 мг/м2, что соответствует 8 кг/га. Тодовая продуктивность раковинных амеб в 40—100 раз выше их биомассы, что связано с небольшой (3—8 дней) продолжительностью жизни одного поколения.
Практическое значение.
Можно предположить, что организмы, встречающиеся в таких количествах, как Arcellinida, могут играть весьма существенную биологическую роль. Раковинные амебы, питающиеся бактериями, водорослями и некоторыми простейшими, сами могут являться добычей некоторых инфузорий, малощетинковых червей, тихоходок, нематод и мальков рыб. Весьма вероятно, что почвенные раковинные амебы участвуют в процессе разложения опада. Неоднократно высказывалось предположение, что эти организмы, отмирая, могут обогащать почву азотистыми соединениями.Выделение фаунистических комплексов раковинных амеб, характерных для определенных условий обитания, делает возможным использование этой группы протистов, довольно хорошо сохраняющихся в субфоссильном состоянии, для историографического анализа изменений этих условий.
Исследование видового состава раковинных амеб в колонках торфа в первую очередь позволяет выделять горизонты, соответствующие различной степени обводнения. Вместе с пыльцевым анализом такой анализ может быть использован для датировки периодов обводнения и осушения в послеледниковый период. Раковинные амебы могут быть использованы и в палеолимнологическом анализе, так как выявление определенных комплексов видов в озерных отложениях дает возможность судить о характере водоема (олиго-, мезо- или эвтрофный), его глубине и наличии или отсутствии плавающей или погруженной растительности в момент образования соответствующего слоя отложений. В последнее время делаются попытки использования раковинных амеб в биодиагностике почв — их видовой состав и соотношение различных морфологических типов раковин являются достаточно чуткими индикаторами физико-химических свойств исследуемой почвы.
Книги:
 Протисты. Руководство по зоологии. Часть 1. Алимов А.Ф. 2000
Протисты. Руководство по зоологии. Часть 1. Алимов А.Ф. 2000
 Протисты. Руководство по зоологии. Часть 2. Алимов А.Ф. 2007
Протисты. Руководство по зоологии. Часть 2. Алимов А.Ф. 2007
 Протисты. Руководство по зоологии. Часть 3. Алимов А.Ф. 2011
Протисты. Руководство по зоологии. Часть 3. Алимов А.Ф. 2011
 Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987
Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987



 Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
 Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423
Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423
 Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989
Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989