


Описание:
К кумовым ракообразным близки танаидовые. Этих рачков не всегда легко заметить из-за их небольших размеров и скрытного образа жизни. В среднем длина их тела составляет 2—3 мм, но многие виды имеют длину всего 1,0—1,5 мм, представителям некоторых семейств свойственны более крупные размеры (10—20 мм). Сравнительно недавно в Филиппинском глубоководном желобе и к юго-востоку от острова Минданао (Филиппины) были обнаружены своего рода гиганты для этой группы животных — Gigantapseudes adac-tylus, достигающий длины 37 мм, и G. maximus — даже 75 мм.
Танаидовые широко распространены в морях и океанах, и лишь несколько видов проникли в солоноватые и пресные водоемы. Всего описано около 800 видов клешненосных осликов.
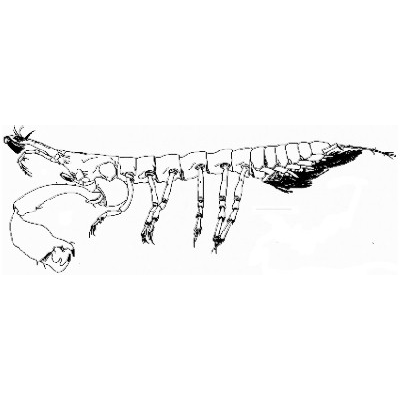
Тело рачков обычно вытянутое, более или менее цилиндрическое либо уплощенное в спинно-брюшном направлении. Короткий карапакс покрывает голову и слившиеся с ней два первых грудных сегмента. Между его боковыми частями и стенкой тела, так же как у кумовых, находятся дыхательные полости. Передний край карапакса обычно вытянут в рострум. Глаза без стебельков — сидячие, с небольшим числом фасеток, расположены на особых глазных лопастях. Эти лопасти неподвижны, не сочленены с головой и тем существенно отличаются от глазных стебельков тонкопанцирных ракообразных, мизид и десятиногих раков. Глаза хорошо развиты у мелководных видов (но далеко не у всех), среди глубоководных же форм виды с глазами очень редки. Обе пары антенн вооружены чувствующими щетинками и несут осязательную функцию. Ротовые части (жва-лы и две пары челюстей) обычно хорошо развиты у самок, у самцов же многих видов они частично или даже полностью редуцируются. Такие самцы не питаются и после копуляции вскоре погибают.
Конечности первого грудного сегмента, слившегося с головой, преобразованы в ногочелюсти. Их крупные эпиподиты выполняют дыхательную функцию (своим движением создают ток воды в дыхательных полостях). Газообмен осуществляется через тонкие покровы эпиподитов, вакуолизиро-ванную ткань боковых частей карапакса и, возможно, через стенки тела, прилегающие к дыхательным полостям. Конечности второго грудного сегмента, также слившегося с головой, крупнее остальных и вооружены настоящими клешнями, за что их называют хелипедами (от греч. chela — клешня и лат. pes — нога), а самих танаидовых — клешненосными осликами. У некоторых видов родов Pagurapseudes и Synapseudes хелипеды самцов асимметричные. Остальные грудные ноги у большинства видов одинакового строения, ходильного типа и служат для передвижения по грунту. Но у ряда форм ноги третьего (первого свободного) сегмента значительно крупнее остальных, с сильно расширенными и укороченными двумя предпоследними члениками, несущими ряды шиповидных игл. Конечности такого строения служат рачкам для закапывания в грунт. Иногда (у самцов Sphy-rapus) длина таких копательных ног достигает длины тела. У рачков, живущих в трубках, три последние пары грудных ног или все, кроме первой, более массивные. Передняя пара (или 3 пары) ног вытянута вперед, а остальные лежат в плоскости, перпендикулярной продольной оси тела. В этой плоскости ноги изогнуты в виде буквы vf так что их «коленкц» и коготки, направленные назад, упираются в стенки трубки. Считается, что такое положение ног позволяет рачкам быстро и ловко передвигаться внутри их домика. Хелит педы и ноги третьего грудного сегмента у ряда видов с редуцированными наружными ветвями — экзоподитами. -
Брюшной отдел (плеон) обычно значительно короче грудного, лишь в редких случаях достигает значительной длины. У Nematotanais mirabilis, например, брюшной отдел лишь в 1,3 раза короче грудного, а у Filitanais rebainsi почти равен ему. Как правило, 5 (реже меньше) передних его сегментов свободные, а шестой срастается с тельсо-ном, образуя так называемый плеотельсон. У ряда видов число свободных сегментов брюшка может уменьшаться вплоть до единственного сегмента у представителей рода Curtipleon. Брюшные ножки — плеоподы (в большинстве случаев в числе 5 пар, редко меньше) — листовидные, как правило, двуветвистые и несут часто длинные' опушенные волосками щетинки, длина которых может превышать длину самих ветвей. С помощью этих ножек танаидовые могут непродолжительное время плавать над дном, и известны случаи, когда они попадались в ловах планктонных сетей. У самцов плеоподы обычно развиты лучше, чем у самок. Самки некоторых видов лишены плеопод. Есть и такие виды, у которых брюшные ножки отсутствуют у обоих полов. К плеотельсону прикрепляется последняя пара ног — уроподы. Они могут быть палочковидными, короткими и малочлени-ковыми или нитевидными, длинными и многочле-никовыми, одноветвистыми или двуветвистыми.
Среди танаидовых есть виды, передвигающиеся по дну с помощью ходильных ног, как например все представители глубоководного семейства Neo-tanaidae. Известна большая группа видов, роющих норки в грунте. К семейству Pagurapseudidae относятся клешненосные ослики, которые, подобно ракам-отшельникам, прячут свое брюшко в пустые раковины брюхоногих моллюсков. Много клешненосных осликов обитает среди зарослей водорослей, поселений гидроидов и губок. Особенно богаты танаидовыми коралловые рифы. Клешненосных осликов находят на разных видах кораллов. Например, Synapseudes асгорогае, как это видно из его видового названия, часто поселяется на широко распространенных кораллах рода Асгорога. Некоторые виды встречаются как на кораллах, так и в коралловом песке (S. menziesi).
Для кораллобионтов характерны очень мелкие размеры, огромные клешни хелипед (часто асимметричные у самцов), необычайно крепкие и, по-видимому, очень цепкие коготки грудных ног и укороченный брюшной отдел. В отличие от большинства депигментированных танаидовых, у клешненосных осликов — кораллобионтов на теле и его придатках обычно имеются многочисленные пигментные пятна и полосы, чаще коричневых, красноватых и фиолетовых тонов. Глаза довольно крупные, коричневого, черного или ярко-красного цвета. У обитающих на рифах Танзании Indoap-seudes brycesoni тело фарфорово-белое, глаза коричневые, а антенны золотистые, у другого вида — S. carinatoides спинная поверхность тела коричневатая, антенны и уроподы с коричневыми пятнами, основания грудных ног зеленого цвета с металлическим отливом, а коготки красные.
На рифах восточного побережья Африки найдены 3 наиболее специализированных вида танаи-довых — кораллобионтов: Tanzanapseudes longi-seta, Т. langi и Т. elegans. Тело этих рачков сильно уплощено в спинно-брюшном направлении, форма его необычна для танаидовых — сферическая или овальная, с 31—32 радиальными выростами, несущими длинные опушенные щетинки, и, как отмечает проф. М. Б э ч е с к у, напоминает звезду. Видимо, такая форма тела способствует лучшему использованию разного рода углублений и ямок в коралловом полипняке, в которые животные прячутся, а наличие многочисленных выростов со щетинками, покрытыми посторонними частицами, хорошо их маскирует.
Среди танаидовых многие виды строят защитные трубки. Длина трубки может в 4—5 раз превышать длину тела животного. Внутренняя поверхность трубки состоит из кутикулярных остатков, соединенных с фекалиями; наружная может быть построена из разного материала: из песчинок, частиц ила, диатомовых водорослей, фрагментов макрофитов (многоклеточных водорослей), раковинок фораминифер и двустворчатых моллюсков. «Строительный материал» ракам приходится выбирать из того, что оказывается у них под рукой в их микробиотопе (местообитании). При построении трубок танаидовые пользуются хелипедами, первой парой грудных ног, а часто и щупиком ногочелюстей. В 1982 г. Джонсону и Аттромандалю удалось наблюдать в аквариумах, как строит трубки Tanais grimaldii. Этих рачков собирали на скалистой литорали близ Бергена, среди зарослей известковой водоросли Corallina officinalis, и помещали в аквариумы вместе с аккуратно вырезанными кусками грунта с водорослями. При постройке трубок животные отламывали куски водорослей мощными хелипедами и переносили их на нужное место, а затем быстро надавливали на них поочередно кончиками грудных ног первой пары, в результате чего кусочки оказывались прочно склеенными. Дело в том, что в покровах грудного отдела тела танаидовых имеются особые железы, слизистый секрет которых выделяется через поры на концевых члениках последней пары грудных ног, за что эти конечности называют прядильными. Секрет выделяется в виде очень тонкой, невидимой невооруженным глазом нити, а пору удалось рассмотреть лишь под электронным микроскопом. С помощью этой же нити рачки могут прикреплять свои трубки к субстрату, причем настолько прочно, что даже сильное течение не может их оторвать. Поврежденные трубки танаидовые быстро ремонтируют. Трубки рачков, живущих в верхнем горизонте литорали, состоят в основном из кусков кораллины и совсем незаметны среди зарослей этой водоросли. У животных, обитающих в верхнем горизонте, подвергающемся длительной осушке и лишенном ковра водорослей, трубки толстые, серого цвета, что также делает их незаметными на фоне окружающего субстрата.
Рачки редко покидают свои «дома», самцы в основном с целью поиска самки («путешествующие самцы»). Трубки у самок более крепкие и длинные и более тщательно построены, чем у самцов. Но как бы небрежно ни была построена трубка у самца Leptochelia savignyi, он самоотверженно защищает свой «дом» от соперников. При приближении другого самца того же вида владелец дома начинает раскачивать переднюю часть тела из стороны в сторону, вытянув при этом вперед хе-липеды с раскрытыми клешнями и тем самым демонстрируя свою силу. Но если эти позы устрашения не пугают пришельца и тот все-таки приближается к входу в трубку, владелец ее может схватить нападающего одной или обеими хелипедами поперек тела и приподнять его над своим ка-рапаксом. В таком положении он некоторое время раскачивает врага, а затем отбрасывает его в сторону. Такие битвы могут продолжаться в течение 5—16 мин, при этом никаких увечий самцы друг другу не причиняют.
Большинство видов танаидовых питается детритом, но есть и всеядные виды. Tanais grimaldii, наряду с диатомовыми водорослями и микроскопическими эпифитами Corallina officinalis, используют в пищу даже кусочки этой известковой водоросли. Было высказано предположение о том, что таким образом рачки удовлетворяют потребности в кальции (хелипеды и карапакс сильно кальцинированы). Одна группа танаидовых (семейство Kalliapseudidae) — фильтраторы. У них на хелипедах и ротовых частях имеется много длинных, густо сидящих перистых щетинок, образующих сложный фильтр для улавливания взвешенных в воде пищевых частиц.
Большинство танаидовых раздельнополы, но для ряда видов известен потенциальный гермафродитизм: один и тот же вид может быть и раздельнополым, и гермафродитным. Например, у Sinelobus stanfordi гермафродитизм наблюдается только в некоторых участках его обширного ареала, в частности на Курильских островах, в то время как в других местах обитают нормальные самцы и самки этого вида. Следует отметить и такой интересный факт, как наличие гермафродитизма двух равных типов — в виде протогинии (явления редкого для животного царства) и в виде протероанд-рии. При протогинии сначала достигающая половозрел ости молодь превращается в самок, которые затем становятся самцами, а при протероандрии — наоборот (сначала самец, затем самка). Аквари-альные наблюдения показали, что при совместном содержании самок и самцов Heterotanais stanfor-di смены пола не происходит, при содержании ювенильной особи вместе с самцом из нее получается самка, если ювенильную особь подсадить к самке/ то из нее вырастает самец, если же молодую особь содержать в одиночестве, то она превращается в самку.
У живущих в трубках Heterotanais oerstedi самец для оплодотворения самки проникает в ее трубку. Самка сначала пытается его вытолкнуть, но затем отступает в глубь трубки, и начинается своеобразный брачный танец, продолжающийся в течение 11 ч. При этом рачки то располагаются «лицом к лицу» и ощупывают друг друга антеннами, то поворачиваются задними концами друг к другу и соприкасаются уроподами. Для принятия таких поз рачкам приходится по нескольку раз переворачиваться через голову («делать кульбиты»). Наконец самец оказывается лежащим на спине, а самка — над ним. Происходит спаривание, и сперматозоиды вводятся в яйцевый мешок, который самка немедленно заполняет яйцами, а ватем начинает массировать его грудными ножками, чтобы яйца и сперма распределились равномерно. Самец через несколько часов после спаривания покидает трубку, самка же плотно запечатывает входное отверстие.
Число откладываемых яиц у танаидовых невелико — у мелких видов обычно от 3 до 10, у крупного Apseudes latrellei отмечено 60 яиц. Развитие прямое, без пелагической личинки. Из яиц, как у кумовых и равноногих ракообразных, вылупляется личинка манка-I. Личинки на этой стадии неподвижны, лишены последней пары грудных ног и плеопод. После линьки они переходят в следующую стадию — манка-II, имеющую зачаточные конечности последней пары и недоразвитые плеоподы. Затем следует ряд линеки после каждой наступает следующая стадия. Обычно различают две ювенильные и две приготовительные стадии и, наконец, рачки переходят во взрослую половозрелую стадию. Но вернемся к стадии манка-II. Личинки на этой стадии готовы покинуть выводковую сумку. У Tanais dulongii самка в последний день перед освобождением личинок из яйцевого мешка вводит в него через гонопору вещество желтого цвета, которое Джонсон и Аттромандаль рассматривают как желток. Им заполняется примерно третья часть мешка, и личинки немедленно начинают его поедать. Уже через несколько часов их прозрачные кишечники становятся желтого цвета. Снабжение личинок желтком перед выходом из яйцевого мешка — явление необычное и неизвестное для других ракообразных. Самка к этому времени уже закупорила вход в трубку, перестала питаться и приготовила для личинок «детскую комнату». Строительство «детской» происходит следующим образом: с помощью тех же нитей, которые используются при построении трубок, самка плетет сетки и ими отгораживает центральную часть трубки. Личинки протыкают выводковую сумку хелипедами и скапливаются в «детской комнате». Самка регулярно инспектирует стенки «детской» антеннами и, в случае необходимости, укрепляет их. Поскольку сети, перегораживающие трубку, засоряются, самка время от времени чистит их, двигая уроподами в спинно-брюшном направлении; при этом их многочисленные щетинки выполняют роль щеток t а ток воды, создаваемый плеоподами, помогает вымывать посторонние частицы. Первой пищей личинкам (после желтка) служат бактерии, детрит и фекалии самки, собираемые с внутренней поверхности материнской трубки. После короткого периода питания личинки покидают трубку, прорывая отверстие в ее стенке хелипедами. Юве-нильные особи начинают строить собственные «дома», используя наружную поверхность материнской трубки как субстрат. Самка производит ремонт стенок своей трубки, разрывает поперечные сети, открывает входное отверстие и начинает снова питаться.
У танаидовых часто хорошо выражен половой диморфизм. Самцы обычно отличаются более мощными клешнями хелипед, более длинными антеннами с большим числом щетинок, иногда иной формой карапакса, грудных сегментов и плеотельсо-на, более развитыми плеоподами и т. п. У самок перед периодом размножения формируется уже упомянутая выше выводковая сумка, обычно состоящая из 4 пар пластинок (реже из одной), отходящих от основания первых четырех пар (или пятой) грудных ног. Иногда имеется пятая пара пластинок, прикрепляющаяся к основанию хелипед. Численность самок, как правило, значительно превышает таковую самцов.
Продолжительность жизни клешненосных осликов невелика: самки Pagurapseudes burgoensis живут до 15 месяцев, а самцы и того меньше — до 9 месяцев.
Tanaidacea широко распространены в морях и оке&нах и встречаются от Арктики до Антарктики. Они обнаружены на всех глубинах — от приливо-отливной зоны до предельных океанических глубин, являются характерным компонентом фауны многих глубоководных желобов. Как уже отмечалось, пресноводных форм средп этой группы ракообразных очень мало, все они безусловно морского происхождения.
Многим видам свойствен огромный диапазон вертикального распределения. Некоторые из таких видов встречаются начиная с верхней сублиторали и кончая ультраабиссалью, например, Leptognathia armata найдена в диапазоне от 80 до 8006 м.
Танаидовые используются в пищу более крупными беспозвоночными, а также рыбами. На мелководье они могут образовывать большие скопления. Например, на илисто-песчанпстой литорали пролива Пьюджет-Саунд (США) средняя плотность поселения Leptochelia savignyi достигает 52 400 экз/м2. Б р а й т отмечает, что в пище глубоководных рыб из района Миссисипи—Алабама— Флорида танаидовые по изобилию, среди прочих ракообразных, занимают третье место.
Книги:
 Жизнь животных. Том 2. Моллюски. Иглокожие. Членистоногие / В. Е. Соколов. — 1988
Жизнь животных. Том 2. Моллюски. Иглокожие. Членистоногие / В. Е. Соколов. — 1988
 Зоология беспозвоночных. От артропод до иглокожих и хордовых. / Вестхайде В., Ригер Р. — 2008
Зоология беспозвоночных. От артропод до иглокожих и хордовых. / Вестхайде В., Ригер Р. — 2008



 Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008