Описание:
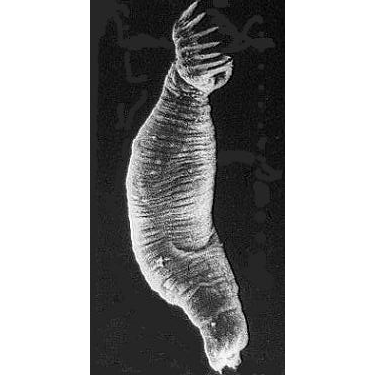
Общая характеристика. Класс моногеней представлен исключительно паразитическими животными. Большинство видов — эктопаразиты (поселяются на наружной поверхности тела животных-хозяев). Моногеней паразитируют главным образом на рыбах, но есть виды, встречающиеся на амфибиях, а один вид — Oculotrema hippopotami — приспособился к обитанию под веками глаз бегемота. Размеры взрослых моногеней варьируют от нескольких долей миллиметра до 3 см.
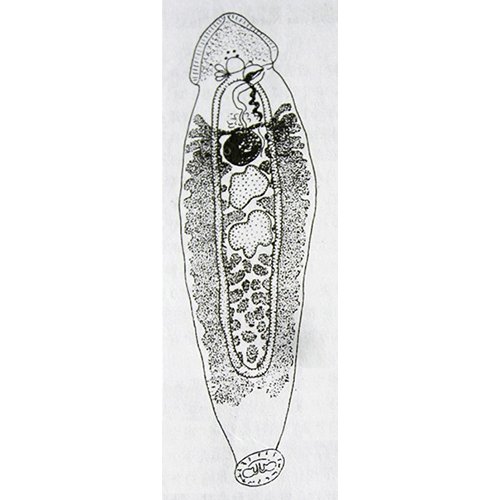
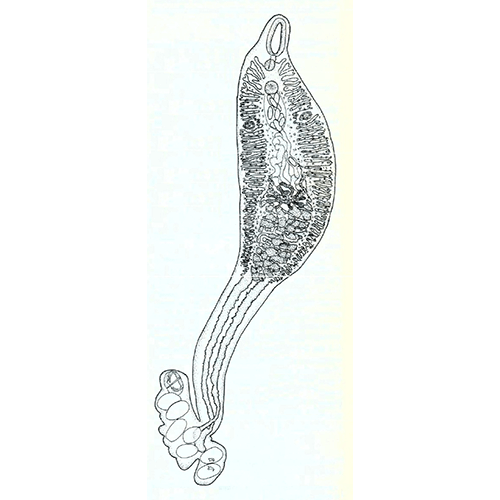
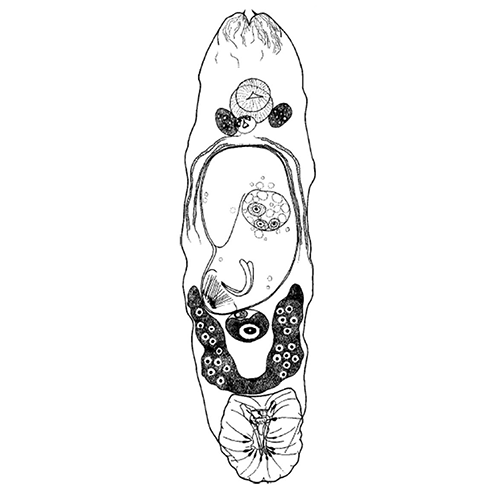
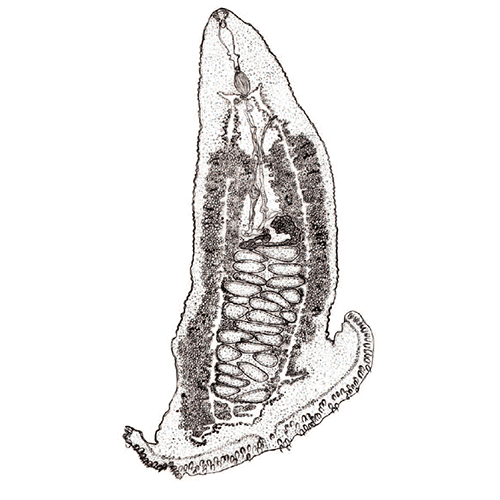
Паразитический образ жизни наложил глубокий отпечаток на строение и биологию моногеней. Тело их отчетливо подразделяется на два отдела: собственно тело, в котором сосредоточены шт внутренние органы, и хорошо развитый прикрепительный диск. Это сложно устроенная образование, которое несет хитиноидные крючке, присоски и специфические для моногеней органы прикрепления — клапаны.
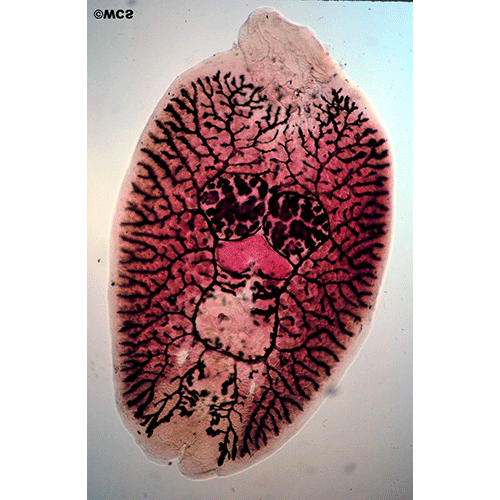
Внутреннее устройство моногеней, хотя и имеет ряд своеобразных черт, в основном соответвует общему плану строения типа плоских червей. Покровы представлены погруженным эпителием (теломентом). Промежутки между органами заполнены таренхимой, нервная система построена по типу этогона, но, в отличие от трематод, имеет развитую поверхностную нервную сеть. Выводные каналы протонефридиев не сливаются, как у трематод, а открываются наружу самостоятельными отверстиями, которые расположены не на заднем, как у других плоских червей, а на переднем конце тела. Кишечник может быть мешковидным, двухветвистым или имеет форму замкнутого кольца. У крупных форм он обильно разветвлен. Ротовое отверстие находится на передам конце. Внутри ротовой полости нередко равзвиваются небольшие присоски, обеспечивающие прикрепление переднего конца тела червя время питания. Этой же цели служат и небольшие выросты, которые несут многочисленные железы, выделяющие клейкий секрет. У многих видов имеется одна или две пары глаз. Половая система гермафродитна. Имеется один, несколько или множество семенников, от которых отходят семепроводы. Сливаясь между собой, они образует толстый, а иногда и длинный семенной пузырек. Женская половая система устроена почти так же, как у трематод, но отличается тем, что имеет особые протоки — влагалища, по которым семя другой особи поступает в семеприемник. Избытки семени и желточных клеток удаляются из оотипа не наружу, как у трематод, а в кишечник, для чего служит специальный кишечно-половой канал.
Органы прикрепления моногеней заслуживают особого рассмотрения. Наиболее распространены кутикулярные крючья, состоящие из прямой рукоятки и сильного серповидного острия. У низших моногеней по периметру прикрепительного диска обычно располагается 10— 16 мелких крючочков, в центре же находится дна (например, у Gyrodactylus) или две (у Tetraonchus) пары крупных и сильных крючьев, приводимых в движение специальными мышцами. Крупные крючья соединены кутикулярными пластиками, к которым крепятся мышцы и которые служат опорой для всего диска. Вонзая крючья в жабры рыбы, паразит плотно заякоривается на хозяине. Благодаря подвижности крючьев моногенеи могут открепляться и переползать с места за место, а иногда и с одного хозяина на другого. Но в некоторых случаях (например, у Ancylodisoides) прикрепление необратимо. Прикрепительный диск обрастает тканями хозяина, и паразит остается связанным с ним на всю жизнь. Для прикрепления к плотным и гладким поверхностям например, к поверхности тела рыбы, обычно служат присоски. В редких случаях (как у паразита осетров Nitzschia sturionis) весь прикрепительный диск превращается в одну большую присоску. Чаще на диске развиваются шесть крупных мускулистых присосок (у Polystoma и др.). Нередко крючья встречаются в сочетании с присосками.
Наиболее совершенные органы прикрепления моногеней — клапаны. Эти органы возникают на основе присосок, укрепленных кутикулярным каркасом. Они действуют наподобие капкана и зажимают между створками жаберные лепестки или участки мягких тканей хозяина. Обычно клапаны располагаются на прикрепительном диске в 2 ряда. У моногеней встречаются и совсем особые способы прикрепления. Так, иногда несущий клапаны диск образует выросты, приобретая форму руки, охватывающей жаберный лепесток. Тропическая моногенея Caballeraxine chainanica обвивает телом жаберный лепесток рыбы и как бы «завязывает» его узлом. Интересно, что клапаны на диске этого вида развиты слабо.
Моногеней питаются исключительно на теле хозяина. Низшие моногеней поглощают слизь и эпителиальные клетки, высшие почти целиком перешли к питанию кровью. Моногенеям свойственна узкая специфичность по отношению к хозяину. Для большей части видов моногеней характерна приуроченность к одному или нескольким близким видам рыб.
Класс моногеней делится на два подкласса: многокрючковые (Polyonchoinеа) и малокрючковые (Oligonchoinea). Первый подкласс объединяет примитивных моногеней. Сюда относятся семейств a Dасtуlogyri d ае, Gyrodасtуlidae и др., на прикрепительном диске которых имеются мелкие краевые и крупные срединные крючья. Ко второму подклассу (это высшие моногеней) относится ряд семейств, органы прикрепления этих животных представлены главным образом клапанами.
Разделение класса на две большие ветви произошло очень давно, возможно, еще в девоне, т. е. в палеозойскую эру.
Жизненный цикл. Жизненный цикл моногеней проходит, за редкими исключениями, без чередования поколений и смены хозяев. Яйца, как правило, задерживаются в матке червей недолго, и развитие личинки происходит во внешней среде. В большинстве случаев яйца крупные и снабжены ножкой или длинными нитевидными выростами — филаментами. Сцепляясь филаментами, они могут образовывать большие скопления, которые медленно опускаются, планируя в толще воды. Они застревают в зарослях водорослей, оседают на теле рыб или могут быть проглочены ими. У отдельных видов развитие личинки завершается еще до того, как яйца выводятся из матки червей наружу. В таких случаях говорят об яйцеживорождении, потому что личинки выходят из яиц тотчас же после их попадания в воду. Есть и живородящие виды, у которых развитие зародышей происходит не внутри яйцевых скорлупок, а непосредственно в матке.
Сформированные личинки внешне несколько напоминают мирацидиев, но, в отличие от них, имеют ц е р к о-м е р — вооруженный крошечными крючочками задний конец тела. У личинок хорошо развиты глаза, одноклеточные железы, протонеф-ридиил зачаток глотки. Нервная система связана с сенсиллами, особенно многочисленными на переднем конце тела. Сформированные личинки передвигаются благодаря биению ресничек и сокращений кожно-мускульного мешка.
В заражении хозяина важную роль играет поведение личинок. Непосредственно после выхода из яйца личинка движется вперед по прямой, но через некоторое время начинает совершать «поисковые» движения — описывать круги и часто менять направление. Личинки обнаруживают положительные и отрицательные фото- и хемотаксисы. Видимо, важный фактор, воздействующий на вылупление личинок,— освещение. Одни виды выходят на свету, другие в темноте, и это можно рассматривать как приспособления к заражению хозяина. Личинки рода Entobdella выходят при смене света темнотой. По-видимому, и это имеет приспособительное значение. В естественных условиях стимулом к вылуплению может оказаться тень, отбрасываемая плывущей рыбой, в результате личинки вылупляются в момент, «удобный» для заражения хозяина. Личинки паразита скатов Acanthocotyle lobiadi вылупляются из яиц под воздействием слизи хозяина, что в естественных условиях происходит в то время, когда скат ложится на дно. Если поблизости ылодятся яйца, на которые попадает слизь, личинки начинают вылупляться и оседать на тело ската.
У некоторых живородящих моногеней личинки лишены ресничек. По мере созревания они активно выползают и матки червя и прикрепляются на поверхности тела того же хозяина, интенсивность заражения которого, таким образом, значительно увеличивается. Расселение паразитов происходит вследствие перехода с рыбы на рыбу взрослых моногеней.
Типы жизненных циклов. Наиболее простые жизненные циклы свойственны примитивным моногенеям — дактилоирусам (Dactylogyrus). Эти черви откладывают яйца на протяжении всего года. Развитие происходит во внешней среде. Вылупившиеся личинки оседают на жабрах рыб и претерпевают метаморфоз, в ходе которого утрачиваются ресничные клетки, формируется вооружение прикрепительного диска, тело метет и развиваются внутренние органы. Через 7—9 дней паразит начинает откладывать яйца. Если следить за зараженностью рыб дактилогирусаин на протяжении года, можно убедится в том, что степень заражения в разные сезоны неодинакова. Зимой на рыбах можно найти лишь единичных червей, весной начинается их бурное размножение и к началу лета интенсивность инвазии достигает максимума. К середине лета зараженность опять снижается, хотя и не так резко, как зимой, а к осени (особенно в южных районах) нередко наблюдается еще один (второй) пик размножения паразитов. Такая сезонная динамика заражения связана с воздействием нескольких факторов, прежде всего с температурой воды. Низкие зимние температуры препятствуют размножению червей. Весеннее повышение температуры, напротив, стимулирует их размножение, что и приводит к резкому возрастанию интенсивности инвазии. Казалось бы, летом, по мере дальнейшего повышения температуры, зараженность рыб должна увеличиваться, но этого не происходит. Дело в том, что температура воздействует не только на паразитов, но и на их хозяев. При высокой летней температуре воды улучшается общее состояние организма рыбы, повышается сопротивляемость заболеваниям. У рыб развивается временный иммунитет к заражению дактилогирусами, что тормозит размножение червей и становится причиной летнего спада заражения. К осени иммунитет рыб снова снижается, и если температура воды еще достаточно благоприятна для размножения паразитов, наблюдается очередной подъем их численности. Зимой же интенсивность инвазии очень низкая. Таким образом, температура воды воздействует на паразитов как непосредственно (стимулируя и затормаживая их размножение), так и опосредованно через организм рыбы, когда появление иммунитета к паразитам становится препятствием к их размножению.
Ход жизненного цикла моногеней часто находится в непосредственной зависимости от жизненного цикла их хозяев. У многих высших моногеней размножение приурочено к определенным периодам в жизни рыб, когда вследствие их скученности (например, во время нереста) переход паразитов с одной рыбы на другую и оседание вылупляющихся личинок облегчены. Такой жизненный цикл характерен, в частности, для паразита сельди Mazocraes alosae, который размножается лишь раз в году, во время нереста хозяина.
Зависимость жизненного цикла моногеней от жизненного цикла хозяина еще более выражена у паразитов лягушек — лягушачьей многоустки Polystoma integerrimum. В отличие от дактилогирусов, многоустка начинает откладывать яйца не через несколько дней, а через несколько лет после заражения хозяина. Лягушка заражается на стадии головастика. Ко времени завершения его метаморфоза, когда крошечный лягушонок выходит из водоема на сушу, паразит локализуется в его мочевом пузыре. Лягушка приступает к размножению только на третий год жизни, когда ранней весной отправляется в водоем для икрометания. Одновременно с хозяином откладывают яйца и многоустки. Их личинки по выходе из яиц прикрепляются к жабрам еще совсем маленьких головастиков. Паразиты проделывают свой метаморфоз довольно быстро и откладывают яйца, из которых выходят личинки, успевающие заразить еще одно поколение головастиков. Это возможно потому, что икрометание лягушек растянуто во времени и головастики, развитие которых продолжается 2—3 месяца, держатся в водоемах до середины лета. Судьба второго поколения многоусток совершенно иная. Обычно личинки паразита попадают на жабры головастиков, уже находящихся на довольно поздней стадии развития, и потому не успевают достигнуть взрослого состояния. Жабры головастика атрофируются, и паразиты, вынужденные покинуть место своего поселения, переползают по брюшной стороне его тела к отверстию клоаки, через которое проникают в мочевой пузырь. Таким образом, многоустки, являющиеся у головастиков эктопаразитами, у лягушки становятся эндопаразитами. Растут они медленно, приобретая способность к размножению одновременно с хозяином. Ведь пока лягушка находится на суше, откладка паразитами яиц, которые могут развиваться только в воде, биологически бессмысленна.
Итак, жизненный цикл лягушачьей многоустки характеризуется рядом своеобразных черт. Он связан с нерегулярной сменой поколений, отличающихся своей локализацией на теле хозяина, продолжительностью жизненного цикла и строением. Первая, так называемая жаберная форма живет недолго и весь жизненный цикл осуществляет на жабрах головастика. Она отличается очень мелкими размерами и недоразвитием ряда органов (нет влагалищ, кишечник не разветвлен и др.)- В сущности, эту форму, приступающую к размножению до завершения морфологического развития, можно назвать н е о т е н и-ческой. Вторая форма сначала тоже живет на жабрах головастика, но заканчивает метаморфоз как эндопаразиту в мочевом пузыре лягушки. Черви растут очень долго и достигают в длину 10—15 мм и начинают размножаться на втором-третьем году жизни под воздействием половых гормонов хозяина, стимулирующих начало размножения. Продолжительность жизни этой формы составляет 4—5 лет. Размножение происходит только раз в году, весной. Описанная смена поколений не обязательна для многоустки, и жаберная форма может отсутствовать.
Совершенно особый тип жизненного цикла свойствен моногенеям из рода гиродактилюса (Gyrodactylus). Этим червям присуще живорождение. В матке взрослой особи развивается только одно яйцо. Оно лишено скорлупки и начинает развиваться после оплодотворения. Это яйцо дает начало не личинке, а взрослому организму, не отличающемуся от материнского ничем, кроме размеров. В матке еще не рожденной дочерней особн тоже закладывается одно яйцо, которое, развиваясь без оплодотворения (т. е. партеногенетп-чески), превращается во внучатую особь. В матке последней, тоже без оплодотворения, развивается правнучатая, а в ней еще одна особь, относящаяся уже к пятому поколению. Наконец материнская особь отрождает дочернюю и в ее матке после этого закладывается новое яйцо. Своеобразный жизненный цикл Gyrodactylus представляет собой гетерогонию, т. е. чередование половых поколений — обоеполых (развивающихся с оплодотворением яйца) и партеногенетических.
Моногеней, будучи эктопаразитами, не нуждаются в системе промежуточных хозяев, получившей большое развитие в эволюции трематод. Однако в отдельных случаях и у моногеней появляются промежуточные хозяева, способствующие расселению паразитов. Это свойственно тем моногенеям, которые паразитируют на хищных пелагических рыбах, обитающих в водах открытого океана. Личинки моногеней, встреча которых со специфичным хозяином затруднена, оседают на теле мелких планктоноядных рыбок. На хозяевах, которых можно назвать промежуточными, личинки проходят только первые стадии своего метаморфоза. В дальнейшем они погружаются в ткани хозяина и переходят в покоящуюся стадию. Когда такие зараженные рыбки становятся добычен хищных пелагических рыб, паразиты переходят на своего специфичного хозяина и на нем завершают развитие. Сведения о жизненном цикле этих моногеней еще очень немногочисленны и многое в ходе цикла остается неясным.
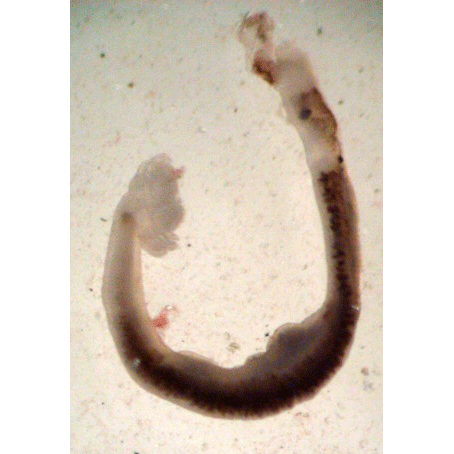
В заключение следует сказать о спайнике парадоксальном (Diplozoon paradoxum), паразитирующем на жабрах карповых рыб. Личинки этих моногеней, попадая на хозяина, могут развиваться и достигать взрослого состояния лишь в том случае, если на рыбе поселяется не одна, а две особи. Личинки сползаются и посредством маленьких присосок крест-накрест соединяются друг с другом. По мере роста и развития червей гкани их в месте соединения срастайся и физиологически они становятся как бы одним животным. Отверстия выводных протоков головой системы сросшихся червей располагаются таким образом, что емя, выделяемое из семепровода од-аой особи, непосредственно поступает во влагалище другой. Таким зеобычным способом у спайника парадоксального всегда обеспечено перекрестное оплодотворение, а значит, и перекомбинация наследственных свойств, столь важная для процветания вида.
Практическое значение. Моногенеи, поселяясь на рыбах, нередко зызывают тяжелые повреждения жабр и других органов и представляют опасность для жизни хозяина, особенно для молоди. Известны многочисленные случаи массовых эпизоотий молоди карпов, вызванных паразитированием моногеней Dactynogyrus vastator. Это теплолюбивый вид, распространенный преимущественно в южных районах. Черви поселяются на жабрах, повреждая их сильными крючьями своего прикрепительного диска. В природных условиях D. vastator распространен широко, но обычно не причиняет хозяину особого вреда. Иначе обстоит дело в рыбоводных хозяйствах. В выростных прудах, относительно неглубоких, хорошо зрогреваемых и густо заселенных выращиваемой молодью, паразиты размножаются особенно интенсивно. Этому способствует и характер жизненного цикла дактилогирусов, которые уже на седьмой день развития приступают к размножению.
В результате на жабрах зараженных мальков насчитывается до 800 червей. Как реакция на заражение возникает уродливое разрастание тканей, жабры разрушаются настолько, что мальки погибают от удушья. В северных районах большой ущерб наносит другой, холодолюбивый вид — Dactylogyrus solidus. В плохо содержащихся хозяйствах, где мальки не получают достаточного питания и слабо растут, паразиты размножаются так бурно, что развивающая молодь рыб гибнет.
Представители рода Gyrodactylus поселяются в основном на коже, жабрах и плавниках рыб. Повреждая своими крючьями покровы, паразиты-моногенеи способствуют попаданию через образовавшиеся ранки бактерий или спор вредных плесневых грибков. Как и дактилогирусы, Gyrodactylus особенно опасны для мальков, интенсивность инвазии которых может достигать 1000 экземпляров на рыбу. При таком высоком заражении начинается обильное отделение слизи, которая вместе с отмершими эпителиальными клетками образует беловатый налет, покрывающий все тело малька. В результате резко снижается уровень кожного дыхания, что в конце концов приводит рыбок к гибели.
Для лечения вызываемых моногенеями заболеваний применяют солевые ванны (5%-ный раствор хлорида натрия в течение 5 мин), вследствие чего паразиты погибают. Для очистительных ванн используют и другие растворы. Очень важно, однако, не допускать массового заражения молоди. Для этого рекомендуется перед заселением прудов обеззараживать их негашеной известью. Особо важное значение имеет правильный уход за выращиваемой молодью, которая в условиях хорошего питания быстро растет и приобретает естественный иммунитет к заражению.
Эволюция моногеней. Проблемы происхождения моногеней и их эволюции подробно разработаны академиком Б. Е. Б ы х о в с к и м (1908—1974). Моногеней, как и трематоды, ведут начало от предковых форм, которые были близки к турбелляриям, но с появлением церкомера обособились от них. Предки моногеней стали паразитами древних палеозойских хрящевых рыб. Первичным местом поселения паразитов были, по-видимому, жабры и жаберные щели. Заражение происходило, скорее всего, пассивно, при затягивании личинок червей вместе с водой в ротовую полость, тесно связанную с жабрами. От моногеней, приспособившихся к паразитированию в ротовой полости рыб, впоследствии возникли паразиты пищевода, желудка и даже кишечника (Diplectanotrema, Enterogyrus и др.). Таким же путем, по мнению академика Б. Е. Быховского, от общих с моногенеями предков могли возникнуть такие специфичные паразиты кишечника, как ленточные черви.
Современные моногенеи — в основном эктопаразиты, но некоторые представители этого класса червей перешли и к эндопаразитизму. Помимо уже упомянутых видов, обитающих в пищеварительном тракте, известны и паразиты других внутренних органов. Это паразиты полости тела рыб, паразиты кровеносной и выделительной систем.Последние две группы явно ведут свое происхождение от жаберных паразитов. Переход от паразитирования на жабрах к кровепаразитизму можно представить себе на примере цикла развития моногеней рода Amphibdella. Личинки этих моногеней, паразитирующих на скатах, легко проникают в кровь через поврежденные жаберные сосуды хозяина. Они интенсивно растут и в период кровепаразитизма становятся самцами (протерандрический гермафродитизм). Позднее у них развивается и женская половая система. Моногеней покидают кровяное русло и возвращаются на жабры хозяина. Смена локализации биологически оправданна. Кровеносная система замкнута, и выход оттуда яиц практически невозможен или требует возникновения специальных адаптаций. Переселяясь на жабры, Amphibdella получает возможность откладывать яйца непосредственно в воду.
Некоторые моногенеи, в прошлом, очевидно, жаберные паразиты, перешли к обитанию в почках, мочеточниках или мочевом пузыре хозяина. Примером возможности такого перехода может быть жизненный цикл лягушачьей многоустки. Нетрудно представить себе возможность подобных случаев и в эволюции моногеней. Причина странной, на первый взгляд, связи между паразитированием на жабрах и в органах выделения кроется в том, что жабры в какой-то мере выполняют и выделительную функцию. Характерно, что среди паразитов разных систематических групп (от простейших до червей) встречаются виды, приспособленные к паразитированию как на жабрах, так и в органах выделительной системы рыб.
Говоря об эволюции моногеней, можно наметить несколько направлений: усложнение и совер шенствование устройства органов прикрепления, формирование строгой зависимости хода жизненного цикла паразита от жизненного цикла хозяина, появление тенденции к живорождению наконец, переход к эндопаразитизму. Однако два последних направления не являются главными в эволюции группы в целом.
Моногенеи, или моногенетические сосальщики (лат. Monogenea) — класс паразитических плоских червей (Plathelminthes). На заднем конце тела взрослых червей расположен характерный прикрепительный диск — гаптор. В роли хозяев для представителей большинства видов выступают рыбы, реже — земноводные и рептилии. Жизненный цикл в подавляющем большинстве случаев включает одно поколение, развивающееся без смены хозяев. Заражение, за редкими исключениями, осуществляет свободноплавающая ресничная личинка — онкомирацидий. Известно около 2 000 видов. Некоторые моногенеи (например, Gyrodactylus и Dactylogyrus) способны приводить к замору рыб в естественных водоёмах и прудах рыбных хозяйств / (Википедия)
Книги:
 Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987
Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987



 Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
 Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423
Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423
 Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989
Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989