































































Описание:
Из всех ракообразных десятиногие издавна пользуются наиболее широкой известностью. Популярный представитель этого отряда — речной рак — неизменный герой бесчисленных сказок и басен. Он фигурирует в числе знаков Зодиака, а с крабами связаны многие мифы инков, ацтеков и древних египтян. К десятиногим принадлежат крупнейшие из ракообразных; длина омаров иногда превосходит 80 см, расстояние между когтями вытянутых в стороны средних ног японского краба Macrocheira составляет 3 м. Благодаря крупным размерам и отличпым вкусовым качествам некоторые десятиногие ракообразные служат важным объектом промысла. В 1983 г. во всем мире было выловлено около 3,2 млн. т ракообразных — креветок, крабов, омаров, лангустов и т. д., т. е. приблизительно в 3,5 раза больше, чем всех лососевых.
Десятиногие ракообразные распространены чрезвычайно широко. Они населяют все океаны и моря, от уреза воды до глубины около 5 км. Особенно богата видами десятиногих тропическая мелководная фауна. В пресных водах обитают общеизвестные речные раки, а в тропических и субтропических странах — крабы и креветки. Наконец, в тропиках обычны сухопутные десятиногие крабы п сильно видоизмененные раки-отшельники.
По образу жизни и по внешнему виду десятиногие весьма разнообразны. К этому отряду принадлежат плавающие креветки, с длинным мускулистым брюшком, ползающие по дну крабы, брюшко которых подвернуто под грудь, раки-отшельники, прячущие свое спирально извитое брюшко в пустые раковины брюхоногих моллюсков, также ползающие по дну речные раки, омары и лангусты, обладающие подобно креветкам длинным симметричным брюшком, и ~ некоторые другие. Общее число видов десятиногих достигает 8500; их приблизительно столько же, сколько видов птиц.
Что же заставляет объединять столь различных животных в один отряд? Всем им свойственны некоторые общие признаки, свидетельствующие об их тесном родстве и едином происхождении.
У всех десятиногих ракообразных три передних грудных сегмента срастаются с головой, а их конечности превращены в ногочелюсти. Пять пар задних грудных вот1 принадлежащих к свободным грудным сегментам, служат для передвижения, почему весь отряд и получил название десятиногих. Часто на некоторых из этих ног есть клешни или иногда подклешни. Жабры целиком покрыты боковыми краями карапакса и снаружи не видны. Самки вынашивают яйца, прикрепляя их к брюшным конечностям, и только самые примитивные креветки просто откладывают яйца в воду. Из яиц выходят планктонные личинки, совсем непохожие на взрослых, у многих пресноводных и глубоководных видов развитие прямое.
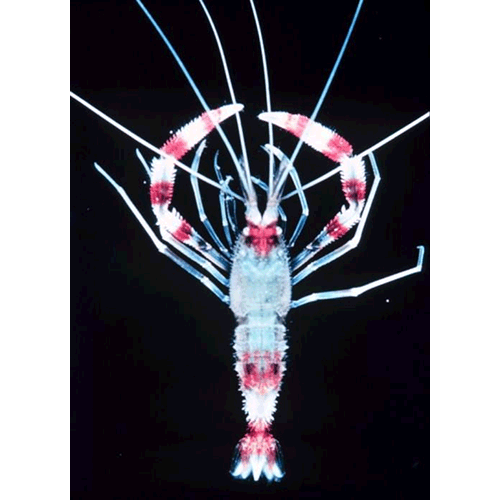
Голова и грудь десятиногих покрыты карапак-сом, плоским и коротким у крабов и цилиндрическим, удлиненным у остальных представителей отряда. Передний конец карапакса часто вытянут в шиповидный, острый на конце рострум. Поверхность карапакса иногда снабжена килями, буграми или шипами, имеющими защитное значение. По бокам от основания рострума прикрепляются стебельчатые глаза. Они могут поворачиваться в разные стороны и обеспечивают широкий кругозор.
Каждый глаз состоит обычно из большого (у креветки Palaemon serratus — 3020) количества сеток, чцсло которых увеличивается с возрастом. Фасетки отделены друг от друга пигментными клетками, и каждая фасетка воспринимает лишь лучи, падающие перпендикулярно на ее роговицу. Она водит только небольшую часть объекта, на который смотрит рак, а другие фасетки видят иные части этого объекта. Таким образом осуществляется «мозаичное зрение». Ночью пигмент расходится к концу и к основанию глаза, и косые лучи могут достигать сетчатки —рак видит объект целиком, но довольно туманно. В воде десятиногие видят только близко расположенные предметы. Рак Galathea на расстоянии 1 Va м не замечает даже крупные движущиеся объекты, но на расстоянии 40 см хорошо различает белые квадраты площадью 2 см2. Поэтому водные десятиногие в поисках пищи, самок, убежища пользуются не столько зрением, сколько обонянием, осязанием и химическим чувством. Зато в жизни сухопутных видов, обладающих особенно длинными глазными стебельками, зрение играет очень важную роль.
В глазном стебельке доводится несколько органов внутренней секреции. Их выделяющиеся в кровь гормоны регулируют расположение пигмента в пигментных клетках, процесс линьки, обмен веществ, содержание в крови сахара и кальция и взаимодействуют с другими органами внутренней секреции.
Передние и задние антенны у крабов очень короткие* а у остальных десятиногих длинные, нередко, особенно у глубоководных креветок, превосходящие по длине тело. У многих «длиннохвостых», т. е. обладающих длинным брюшком, деся-сятиногих на каждой из передних антенн находится по два или даже по три жгута. Эти жгуты богаты чувствительными щетинками и цилиндрами, выполняющими функции органов обоняния, осязания и химического чувства. В основном членике каждой из передних антенн расположен орган равновесия — статоцист.
У большинства десятиногих полость статоциста соединяется с наружной средой узкой щелью. На стенках полости находятся чувствительные щетинки двух типов — крючковидные и нитевидные. На крючковидные щетинки опираются один или несколько статолитов. Большинство десятиногих использует в качестве статолитов песчинки, которые после линьки рак запихивает в полость статоциста, у некоторых статоцист образуется за счет затвердевания выделяемого специальными железами вещества; у многих крабов статолитов нет. Ощущая давление статолита на те или иные чувствительные щетинки, животное ориентируется по отношению к силе земного притяжения.
Для выяснения механизма действия статоцистов был проделан такой остроумный опыт. Креветок Palaemon содержали в аквариуме с грунтом из железных опилок. После линьки рачки вынуждены были вложить в свои статоцисты вместо песчинок частицы металла. При приближении сверху к аквариуму магнита креветки дружно поворачивались брюшной стороной вверх* поскольку этого требовали их притягиваемые магцитом железные статол&ты. Поднося магнит к стенке аквариума, можно было легко заставить животных повернуться боком и т. д. Действия нитевидных щетинок не связаны со статолитами. Они регулируют повороты тела десятиногих по отношению к вертикальной оси.
Жвалы десятиногих всегда мощные и служат для перетирания и разрывания пищи. Передние челюсти маленькие, а на задних имеется крупная наружная лопасть — скафогнатит, благодаря движениям котороц осуществляется смена воды в жаберной цолости. Она совершает от 120 до 200 взмахов в 1 мин, причем в жаберной полости в течение 10 мин сменяется от 2 до 25 объемов водц, что зависит от температуры воды1 содержания в ней кислорода и вида рака.
Из 8 пар грудных конечностей 3 передние превращены в ногочелюсти. Они принимают участие в захватывании пищевых частиц и в передаче их к ротовому отверстию. Остальные 5 пар грудных ног служат главным образом для ползания и, кроме того,. передние из них — для хватания пищи. На таких хватательных ногах развиваются клешни. У крабов, раков-отшельников и других близких к последним форм клешни есть только на передней паре ходильных ног, у большинства креветок — на двух передних парах, а у речных раков, омаров и креветок из семейств Penaeidae и Sergestidae — на трех передних парах, причем клешни первой пары значительно крупнее остальных. У креветок из семейства Crangonidae на передних ходильных ногах настоящие клешни заменены подклешнями. У лангустов, раков-медведей и некоторых других клешней нет совсем.
Нередко левая и правая клешни первой пары имеют разное строение: одна из них крупнее или они различны по форме. Например, у омаров одна (то левая, то правая) более массивная клешня приспособлена для раздавливания панцирей морских ежей, раковин моллюсков, домиков морских желудей и носит название дробящей, а другая, более тонкаяf режущая клешня помогает делить на куски тело рыб, креветок и т. д. Пресноводные родственники омаров — речные раки унаследовали от своих предков асимметрию клешней. Особенно резко выражена асимметрия клешней у самцов тропических полусухопутных так называемых манящих крабов, обладающих маленькой левой и колоссальной правой клешней, сгибая которую они производят своеобразные манящие движения. У раков-щелкунов (сем. Alpheidae) одна из двух передних клешней также значительно крупнее другой и устроена таким образом, что подвижный палец может с силой прижиматься неподвижному, причем раздается резкий звук.
Задние грудные нога в некоторых случаях также имеют какие-нибудь^ специальные приспособления. У крабов-плавунцов (сем. Portunidae) их последние членики уплощены, ими крабы отталкиваются от воды, когда плавают. У раков-отшельников задняя пара грудных ног укорочена и служит для удерживания раковин брюхоногих, в которых они живут. Родственные им Lithodidae в процессе эволюции утеряли способность жить в раковине, но сохранили укороченные задние грудные ноги. Эти конечности у Lithodidae подвернуты под край карапакса и используются для очистки жабр.
С грудными конечностями связаны жабры. Они прикрепляются или к основному членику ногоче-люстей и~ ходильных ног, или к сочленению между этим члеником и телом, или, наконец, к стенке тела над местом прикрепления той или иной грудной конечности. Часто на каждом грудном сегменте расположены жабры всех трех иди двух типов. Жабры прикрыты боковым краем карапакса, ограничивающим снаружи жаберную полость.
У плавающих креветок вода свободно проникает в жаберные полости и, направляемая движениями скафогнатита задней челюсти, омывает жабры. У ползающих десятиногих нижние края карапакса плотнее прижимаются к стенке тела, основные члендки ног развиты сильнее и благодаря этому жаберные полости почти полностью изолированы от внешней, среды, что препятствует загрязнению жабр грунтом. Для проникновения воды в жаберные полости имеются специальные от* верстия, расположенные у крабов по одному церед основанием клешней, а у раков-отшельников, речных раков, омаров и других между основания* ми ходильных ног. Из жаберной полости вода выходит из-под нижнего края карапакса в его передней части.
Зарывающийся в грунт краб Corystes складывает свои задние антенны так, что они образуют трубку, которую краб выставляет над поверхно* стью грунта. По этой трубке вода достигает жаберных полостей краба. Близкая к ракам-отшель-никам Albunea имеет сходное приспособление, но ее дыхательная трубка образуется не задними, а передними антеннами. Краб Calappa, зарываясь, разгребает клешнями грунт так, что перед выходами в жаберные полости остается свободное пространство, заполненное водой. Многие закапывающиеся крабы цмеют на всем крае голово-грудного щита от глаз до основания клешней ряд щетинок, действующих как фильтр, мешающий песчинкам попасть в жаберную полость при дыхании.
У многих сухопутпых десятиногих дыхательная система сильно изменяется. На внутренней поверхности стенок жаберных полостей у сухопутных потомков раков-отшельников — пальмового вора (Birgus latro) и Coenobita — развиваются гроздевидные складки кожи, в которых разветвляются многочисленные кровеносные сосуды. - Это — настоящие легкие, позволяющие использовать кислород заполняющего жаберные полости воздуха. Легкие вентилируются благодаря движениям скафогнатита, а также вследствие способности животных время от времени приподнимать и опускать карапакс* для чего служат специальные мышцы. Замечательно, что при этом сохраняются и жабры, правда, относительно малых размеров. Удаление жабр нисколько не вредило дыханию Birgus и Coenobita; с другой стороны, оба рака полностью потеряли способность дышать в воде. Погруженный в воду пальмовый вор погибал через 4 ч. Остаточные жабры, по-видимому, не функционируют.
За грудным отделом тела следует брюшной,: строение которого различно у разных десятиногих и тесно связано с их образом жизни. У плавающих десятиногих — креветок — брюшко сильно развито, длиннее головы и груди, часто сжато с боков и снабжено относительно крупными плавательными ножками — плеоподами. По паре плёопод прикреплено к каждому из пяти передних брюшных сегментов, а конечности последнего брюшного сегмента — уроподы — широкие, пластинчатые и образуют вместе с тельсоном хвостовой веер. Ползающие по дну длиннохвостые ракообразные (омары, речные раки, лангусты и др.) также имеют длинное брюшко, но оно более или менее сжато в спинно-брюшном направлении, а плеоподы относительно короткие и слабые и для плавания не служат. У большинства раков-отшельников покровы брюшка мягкие, кожистые и оно спирально завернуто, так как помещается внутри пустой раковины брюхоногих моллюсков и следует 8а ее оборотами. Асимметрия брюшка раков-отшельников проявляется и в строении его конечностей: плеоподы развиваются только на стороне брюшка, прилегающей к наружной стенке раковины, а уропода с этой стороны гораздо крупнее, чем с противоположной. У некоторых родственных ракам-отшельникам десятиногих, например у представителей семейства Litho-didae, к которому, между прочим, принадлежит камчатский краб, а также у сухопутных пальмового вора и Coenobita брюшко укорочено и подвернуто под грудь, но и у них в расположении хитиновых щитков, покрывающих брюшко, и плеопод заметны признаки асимметрии, свидетельствующие об их происхождении от форм со спирально закрученным брюшком. Наконец, у настоящих крабов брюшко сильно уменьшено в размерах и всегда подвернуто под грудь, число составляющих его сегментов, особенно у самцов, сокращено, плеоподы недоразвиты, а уроподы отсутствуют.
Первичная функция плеопод плавательная, как это свойственно креветкам. Однако даже у креветок плеоподы, кроме того, участвуют в процессе размножения. У самцов креветок первая или вторая пара плеопод частично преобразована в совокупительный орган, а самки, за исключением представителей семейств Penaeidae и Sergestidae, прикрепляют к плеоподам яйца. У ползающих десятиногих половая функция плеопод стала основной, самки используют их только для вынашивания яиц, самцам передние плеоподы служат копу-лятивным органом. У самцов крабов сохраняются только первая или две первые пары плеопод, превращенные в копулятивный аппарат.
Десятиногие — весьма подвижные животные. По способу передвижения их можно разделить на плавающих и ползающих, причем к первым принадлежат почти все креветки, а ко вторым остальные. Резкой границы между этими группами нет. Большинство креветок может не только плавать, но и ползать по дну или растениям, а среди таких типичных ползающих десятиногих, как крабы,: известно обширное семейство крабов-плавунцов (Portunidae), представители которого приспособились к плаванию. Креветки плавают при помощи плеопод, отталкиваясь ими от воды, причем прижимают антенны и грудные ноги к телу и. выпрямляют брюшко. Планктонные виды держатся в воде вертикально, работая не только; плеоподами, до и наружными ветвями грудных ног: При этом они вытягивают в стороны свои длинные антенны и внутренние ветви трудных ног и таким, образом препятствуют погружению. При приближенйи врага креветки резкими скачками уплывают задом наперед, порывисто сгибая брюшко и отталкиваясь тельсоном и уроподами от воды.
Ползание по дну осуществляется при помощи четырех пар задних грудных ног. Десятиногие сгибают и разгибают их последовательно, причем никогда левая и правая ноги одной пары не действуют одновременно. Омары, лангусты, раки-отшельники ползают передним концом вперед, но скорость их передвижения невелика. Гораздо быстрее бегают десятиногие с укороченным грудным отделом и подогнутым под него брюшком, т.е. крабы и крабовидные родичи раков-отшельников— камчатский краб, пальмовый вор, Coenobita и др. Все они передвигаются боком, достигая иногда очень большой скорости. Обычный «травяной краб» Carcinus maenas бегает со скоростью 1 м/с, а сухопутный краб-привидение Ocypode мчится на своих вытянутых ногах так быстро, что умудряется ловить даже мелких птиц. Крабы-плавунцы также плавают боком. При этом вторая — четвертая пары грудных ног делают 630—780 взмахов в 1 мин, а последняя пара работает еще более интенсивно.
Вошедшее в басни и поговорки мнение о том, что «рак пятится назад» не совсем справедливо. В нормальной для него обстановке, на дне водоема, он этого не делает. Когда же пойманного рака выпускают обратно в воду, он начинает, так же как креветки, сгибать брюшко и действительно, плывя толчками задом наперед, быстро опускается на дно.
Окраска десятиногих разнообразна. Большинство донных ползающих видов бурого или зеленоватого цвета, напоминающего цвет грунта, на котором они живут. Обитающие среди водорослей или ползающие по ним креветки и крабы зеленого цвета. Прикрепляющийся к саргассовым водорослям и к другим плавающим на поверхности моря предметам краб Planes minutus окрашен по-разному: особи, сидящие на водорослях, бурые или зеленоватые, а сидящие на голубых хондрофо-рах — синеватые. Очень пестра окраска многочисленных крабов — обитателей коралловых рифов, но и она делает рачков незаметными на фоне многокрасочного кораллового известняка. Тропический сухопутный краб-привидение Ocypode имеет цвет песка. При этом на ярком солнце его тело отбрасывает на светлый песок темную тень. При приближении врага краб подгибает ноги и прижимает плоский карапакс к песку, делаясь совершенно незаметным.
Наряду с этими и многими другими примерами покровительственной окраски десятиногих можно указать и случаи существования у них предупреждающей окраски. Карапакс африканского сухопутного краба Sesarma-meinerti пурпуровый и окаймлен желтым кантом, а клешни ярко-крас-ные. При приближении врага Sesarma не прижимается к песку, как краб-привидение, а, наоборот, приподнимается на своих ходильных ногах и потрясает в воздухе клешнямяг У самцов манящих крабов цвет их крупной клешни обычно резко контрастирует с цветом карапакса, а также грунта, что делает манящие движения клешней еще более заметными. Так, карапакс Uca batuenta золотисто-коричневый, а клешня совершенно белая, у U. beebi карапакс зеленый, большая часть клешни желтая или розовая, а ее пальцы белые.
Все эти типы окраски определяются присутствием в покровах пигментов, большая часть которых представлена каротиноидами. Особенно обычен из них астаксантин, в .чистом виде имеющий красный цвет. Соединяясь с белками, он образует пигменты синеватых и буроватых тонов. При повышении температуры эти соединения разрушаются, астаксантин освобождается. Поэтому раки при варке краснеют.
Помимо десятиногих, _ цвет которых остается неизменным, существуют виды, меняющие свою окраску в зависимости от цвета грунта, на котором они находятся, или в зависимости от времени суток. Креветки Hippolyte, Palaemon и Palaemone-tes на светлом грунте приблизительно через 1У2 ч светлеют, а на темном темнеют. Наряду с этим ночью они светлеют, становясь прозрачными, а днем приобретают ту или иную окраску. Креветка Crangon, наоборот, ночью окрашена темнее, чем днем. Манящие крабы темнеют во время отлива, когда они активны, и светлеют во время прилива, когда они сидят в своих норах.
В толще покровов таких десятиногих расположены многочисленные, снабженные длинными разветвленными отростками органы, называемые хроматофорами и содержащие огромное количество зерен пигмента. У некоторых креветок в каждом хроматофоре находится несколько пигментов, причем каждый из них занимает свои ответвления хроматофора. Хроматофоры Crangon имеют ответвления с черным, белым, желтым и красным пигментами, хроматофоры Penaeus устроены так же, но черный пигмент в них заменен синим. У крабов и других ползающих десятиногих каждый хроматофор содержит один или самое большее два пигмента. Когда все пигментные зерна концентрируются в середине хроматофора, животное светлеет, когда же пигментные зерна распространяются по всем отросткам, оно темнеет. Если в отростках находятся пигментные зерна одного цвета, а остальные собраны в центре хроматофора, креветка оказывается окрашенной в цвет распыленного пигмента.
К хроматофорам не подходят нервы. Как показано многочисленными опытами, их деятельностью управляют органы внутренней секреции, находящиеся главным образом в глазном стебельке, а также в головном ганглии и в отходящих от него окологлоточных нервных тяжах. Световое раздражение, воспринимаемое глазами, передается этим брганам, а они выделяют в кровь различные гормоны. Одни гормоны заставляют зерна того или иного пигмента распространяться по отросткам хроматофора* другие, наоборот, вызывают их концентрацию. Отрезание глаз или тугая перевязка глазных стебельков полностью нарушает движение зерен пигмента в хроматофорах г так как преграждает путь гормонам в кровь.
Замечателен автоматизм движений пигмента в хроматофорах некоторых ракообразных. У манящих крабов эти движения^ как уже указывалось, точно согласованы с приливо-отливным ритмом. У крабов Uca pugnax из двух соседних участков, валивающйхСя приливом из-за разной конфигураций берега в разное время, периодичность изменения окраски различна. Эти крабы вместе помещались в темну*> Комнату, где каждый из них сохранял присущий ему ритм передвижений пигмента в хроматофорах. Круглосуточное освещение или, наоборот, помещение крабов в условия постоянной темноты, а также содержание их при различной температуре (от 6 до 26 °С) не нарушает этого четкого ритма. Учитывая разницу в часах прилива и отлива в разных частях земного шара, крабов Uca pugilatof на самолете перевезли с Атлантического побережья Америки на Тихоокеанское. Невольные путешественники сохранили в новом местообитании выработавшийся у них на родине ритм распыления и концентрации пигментных зерен в хроматофорах. Все это позволяет считать миграции пигмента в хроматофорах манящих крабов одним из лучших примеров существования «биологических часов», идущих точно и правильно независимо от условий внешней среды.
Все же некоторые внешние факторы влияют на хроматофоры манящих крабов. Замечено, например, что при усилении космической радиации степень распыленности темного пигмента в хроматофорах увеличивается; крабы таким образом защищаются от вредного действия коротковолновых излучений. Повышение атмосферного давления уменьшает интенсивность космической радиации, и можно думать, что крабы реагируют на колебания атмосферного давления^ а не непосредственно на излучение. Для проверки этой гипотезы крабов помещали в камеру с постоянным атмосферным давлением, причем окавадось^ что расположение пигмента в их хроматофорах меняется в соответствии с интенсивностью космической радиации независимо от атмосферного давления.
Способы и объекты питания десятиногих довольно разнообразны. Большинство этих ракообраа* ных можно считать хищниками. Морские ползающие виды находят различных животных — поли-хет, моллюсков, иглокожих, других ракообразных, разрывают или раздробляют добычу клешнями, перетирают ее жвалами и поедают. Многие способны расправляться с крупными и сильными жертвами. Краб Loxorhynchus grand is (с карапаксом длиной 26 см и шириной 21 см) разрывал клешнями осьминогов и больших морских звезд. Некоторые десятиногие наряду с животными поедают и богатый органическим веществом грунт, а также растения. Раки-отшельники охотно едят разных беспозвоночных, но наряду с ними и грунт, который они захватывают своей меньшей (обычно левой) клешней. Креветки Pandalus, Palaemon, Crangon и др. также едят главным образом мелких животных, но заглатывают и грунт и водоросли. Наши речные раки питаются преимущественно водными растениями — урутью, роголистником, рдестом, харовыми водорослями, но при случае охотно поедают моллюсков, личинок насекомых, а также трупы любых животных. Чрезвычайно широк ассортимент объектов питания некоторых сухопутных десятиногих. Пальмовый вор ест падающие на землю плоды различных пальм, особенно охотно маслянистые плоды пан* данового дерева, грунт, а также нападает на других сухопутных ракообразных, в том числе на больных особей своего вида. Мнение о питании его кокосовыми орехами неверно. Несмотря на огромную силу своих клешней, он не может разгрызть скорлупу зрелых кокосовых орехов и поедает только разбитые. Помещенный в садок с кокосовыми орехами, пальмовый вор через несколько недель погибал от голода. Неверно также мнение, что пальмовый вор забирается на кокосовые пальмы, чтобы сбросить орехи вниз. Он не может влезать высоко на деревья и совсем не способен слезать с них. Вообще никакой связи между ним и кокосовыми пальмами нет, поскольку он обитает на многих островахг на которых эти пальмы отсутствуют.
Некоторые десятиногие используют в пищу грунт. Тропические и субтропические пресноводные креветки из семейства Atyidae на концах пальцев клешней имеют кисточки щетинок, при помощи которых захватывают ил и передают его в рот. Так же питаются и пещерные представители этого семейства, в том числе живущая в пещерах Закавказья, Балканского полуострова и южной Франции Troglocaris anophthalmia. Полусухо-путные манящие крабы также питаются грунтом, поглощая его в огромном количестве. Вокруг отверстий их нор всегда располагаются комочки их экскрементов, состоящие из пропущенного через кишечник грунта. Морские Galathea метут дно щетинками задних ногочелюстей, причем частицы грунта, разлагающихся водорослей и т. д. застревают между щетинками, а затем счищаются вторыми ногочелюстями и отправляются в рот. Более крупные пищевые частицы раки хватают клешнями.
Фильтраторы среди десятиногих немногочисленны. Живущие в норах Callianassidae энергично машут плеоподами, вызывая таким образом сильный ток воды, направленный в нору. Этот ток проходит через сложенные в виде корзинки передние грудные ноги, густо обсаженные щетинками, которые задерживают взвешенные в воде частицы. Время от времени раки резко сгибают брюшко, и получающийся при этом обратный ток воды промывает фильтрующую камеру, освобождая ее от попавших в нее крупных частиц. Крабовидный рак Porcellana машет задними ногочелюстями, снабженными многочисленными щетинками, отфильтровывая ими мелкие взвешенные частицы.
При добывании пищи основное значение у большинства водных десятиногих имеют чувства обоняния и осязания. После внесения в аквариум с крабами или раками-отшельниками пищи животное реагирует на нее характерными движениями передних антенн, богатых чувствительными цилиндрами, и, почуяв добычу, приступает к ее поискам. Дальнейшее поведение камчатского краба Д. Н. JI о г в и н о в и ч описывает так: «В выборе направления (к пище) краб руководствовался тщательным ощупыванием бетонного дна бассейна концами клешней. Краб опускал клешни вертикально вниз и, касаясь грунта концами пальцев клешней, быстро закрывал и открывал их, постепенно обследуя площадку перед собой. Эти щупающие движения чрезвычайно характерны, энергичны, «нервны». Краб искал как бы вслепую, двигаясь чрезвычайно запутанным маршрутом. По мере приближения к корму краб приходил в большое возбуждение, что выражалось в усилении ощупывающих движений клешней по грунту. Однако даже в непосредственной близости от корма (например, на расстоянии 1 см от конца клешней до корма) краб неоднократно промахивался и удалялся от него. Это указывает на то, что осязание концов клешней не выправлялось обонянием или зрением. Наступает момент, когда положение кормового объекта точно определяется крабом, что выражается в стремительном схватывании добычи, которое производится правой или левой клешней или обеими сразу». В опытах Д. Н. Логвинович из-за неспособности направиться прямо к предлагаемой пище краб терял на путь к ней несообразно много времени. В одном случае краб достиг корма, положенного от него на расстоянии 75 см, через 23 мин, причем исколесил 930 см* в другом случае для достижения корма лежавшего в 260 см, потребовалось всего 17 мин, а в третьем случае, когда корм находился всего в 5 см от краба, последний искал его в течение 65 мин.
Важная роль органов обоняния и осязания при поисках пшци подтверждается й опытами над креветкой Palaemon. После удаления глаз креветки отыскивали пищу в течение 4-^5 мин, но когда у них были удалены и передние антенны, время разыскиваний увеличилось до 20 мин, однако даже лишенные обеих нар антенн креветки в конце концов все-таки находили пищу, чему помогали чувствительные щетинки ротовых придатков и пальцы ходильных ног.
Несколько иначе происходит охота за движущимися объектами, при которой основное значение приобретает зрение. Хищные крабы неподвижно лежат на дне и высматривают добычу. При виде движущейся жертвы они бегут, а крабы-плавунцы быстро плывут прямо к ней и обычно ее настигают. Многие крабы, в особенности крабы-плавунцы, таким образом успешно ловят рыб.
Все сухопутные десятиногие при поисках пищи руководствуются в первую очередь зрением. Пальмовый вор видит пищу с расстояния в несколько метров и прямо направляется к ней. Он успешно преследует других сухопутных десятиногих, например краба Cardisoma. Крабы-привидения (Ocypode) благодаря своим необыкновенно длинным глазным стебелькам и сложно устроенным глазам издали видят добычу и умудряются, как уже указывалось выше, ловить даже птиц.
Будучи в большинстве хищниками, десятиногие сами подвергаются нападению многочисленных врагов — рыб, головоногих моллюсков и других десятиногих, а сухопутные виды — птиц и грызунов. Для защиты от врагов десятиногие выработали разнообразные и совершенные приспособления. К таким приспособлениям принадлежат разобранные ранее примеры покровительственной окраски, свойственной некоторым креветкам и крабам, и угрожающие позы многих крабов. Той же цели служат и другие особенности строения и поведения десятиногих.
Многие ползающие десятиногие, например омары, лангусты, крабы, в том числе обычные в Черном море Pachygrapsus и Carcinus, укрываются между камнями и раковинами моллюсков, в расщелинах скал и т. д. Существуют крабы и креветки, которые, стремясь к безопасности, закапываются в грунт. Креветки Crangon, Nectocrangon и, вероятно, другие представители семейства Crangonidae, а также донные креветки из семейства Penaeidae днем зарываются в грунт и сидят в нем горизонтально, выставив наружу глаза и антенны, а ночью выходят на охоту. Так же поступают крабы-плавунцы, Galatheidae, некоторые раки-отшельники. Многие крабы, зарывшись в грунт» располагают свое тело вертикально.
Различные десятиногие строят более или менее совершенные норы, в которых проводят значительную часть жизни. Представителей семейства Callia-nassidae называют «ракамк«кротами» за то, что они вырывают глубокие ходы, часто имеющие форму латинской буквы U, а иногда и более сложную. Рачок роет грунт клешнями, выбрасывая его из норы задними грудными ногами. Стенки хода обмазываются илом или выделениями кожных желез рачка.
Менее совершенны норы речных раков, которые они вырывают клешнями в откосах берегов рек и ручьев. В этих норах раки проводят дневное время, а кроме того, зимуют. Некоторые виды американских речных раков из рода Cambarus строят целые туннели длиной до 75 см. Расселившийся в недавнее время по берегам морей и рекам Западной Европы китайский мохнаторукий краб Eriocheir sinensis делает норы в откосах берегов низовьев рек, куда еще доходит прилив. Эти норы наклонные и построены так, что в их низко расположенной концевой части остается морская вода после отлива. Е. sinensis своими норами повреждает плотины, оросительные каналы и другие гидротехнические сооружения.
Почти все десятиногие, приспособившиеся к жизни на суше, сооружают норы, имеющие в их жизни большое значение. Обычно норы сухопутных крабов настолько глубоки, что достигают уровня грунтовых вод и их обитатели могут пить находящуюся в их домах воду. В других случаях поры расположены так, что они заливаются во время прилива. Часто норы имеют два выхода. Некоторые манящие крабы, влезая в нору, закрывают вход в нее крышкой из грунта. В каждой норе обитает один краб. Рытье нор осуществляется ходильными ножками одной стороны тела и меньшей клешненосной ногой, причем краб располагается в строящейся норе боком.
Манящие крабы образуют на илистых прибрежных отмелях тропиков густые поселения. На 1 м2 нередко насчитывается 50 и более их нор. При приближении опасности все население такого поселка дружно и молниеносно скрывается в своих убежищах. Крабы замечают летящую птицу на расстоянии 7 м, а человека — на расстоянии 9 м. Они оповещают своих соседей об опасности ударами клешней о грунт. Благодаря колебаниям грунта сигнал принимается даже в том случае^ когда крабы не видят друг друга.
Пальмовый вор выкапывает в почве неглубокие норы, которые выстилает волокнами кокосовых орехов. Ч. Дарвин рассказывает, что туземцы на некоторых островах выбирают из нор пальмового вора эти волокна, нужные им в их немудреном хозяйстве. Иногда пальмовый вор довольствуется естественными убежищами — расщелинами в скалах, полостями в осушенных коралловых рифах, но и в таких случаях использует для их обкладки растительный материал, сохраняющий в жилье повышенную влажность.
Некоторые крабы сами нор не роют, а используют в качестве жилища норы или трубки других животных, например трубки сидячих полихет. В трубках червя Chaetopterus поселяются разные представители семейств крабов Pinnotheridae и крабовидных Porcellanidae. Последние живут также в ходах, выточенных в известняке моллюском-камнеточцем Pholas.
Раки-отшельники используют пустые раковины брюхоногих моллюсков, пряча в них длинное,; лишенное твердых покровов брюшко. Они подбирают себе раковины по размерам и плотно в них удерживаются уроподами и укороченными задними грудными ногами. На врагов рак-отшельник реагирует, забираясь целиком в раковину и закрывая ее устье своими клешнями. Раки-отшельники одного вида могут заселять раковины разных видов моллюсков. Например, Pagurus prideauxi использует раковины не менее 25 видов брюхоногих. Однако раки-отшельники, по-видимому, отдают предпочтение раковинам определенных видов. В аквариуме Pagurus acadianus выбирал в первую очередь раковины Buccinum, во вторую — Thais и в третью — Littorina, что соответствует частоте использования раковин этих видов раком-отшельником в естественных условиях. Предпочтение, оказываемое одним раковинам перед другими, связано с различным отношением внутреннего объема раковины к ее массе. При выборе раковины ракиютшельники руководствуются не зрением, а осязанием. Насильно извлеченные из своего убежища, они стремятся возвратиться в старую раковину, несмотря на возможность занять предлагаемую им другую того же вида и размера. Однако после линьки старая раковина оказывается тесной для ее владельца и он подыскивает новую, более вместительную. Искусственно лишенный раковины рак-отшельник чувствует себя крайне неспокойно, напоминая, по выражению Тэйлорал купальщика, у которого украли одежду.
Некоторые виды раков-отшельников поселяются не в спирально извитых раковинах брюхоногих моллюсков, а в почти прямых конусовидных раковинах лопатоногнх моллюсков или даже в кусках бамбука. У таких видов брюшко вполне симметрично. Близкий к ракам-отшельникам сухопутный рак Coenobita прячет свое мягкое брюшко в пустые раковины наземных моллюсков. Обитающий на Бермудских островах С. diogenes использует в качестве жилья только раковины Li-vona pica. Этот вид здесь вымер, и рак разыскивает его полуископаемые остатки. Других подходящих моллюсков на Бермудских островах нет, и чрезвычайно интересног какая судьба ждет С. diogenes в дальнейшем, когда запас пустых раковин Livona истощится.
От поселения в чужих ходах и пустых раковинах один шаг к жизни на поверхности тела и в полости тела живых животных, которые могут быть также использованы как убежища. Такой шаг многие десятиногие ракообразные сделали.
Существуют креветки, проводящие всю жизнь внутри губок. В двух губках Speciospongia ves-рага общим объемом 50 000 см3 было обнаружено 18 000 экземпляров двух видов раков-щелкунов Synalpheus. Креветки ив рода Spongicola сидят попарно (самец и самка) в полостях стеклянных губок t куда они попадают еще будучи личинками и откуда затем, увеличившись в размерах, выбраться уже не могут. Другие креветки и некоторые крабы обитают под колоколом медуз, есть десятиногие, живущие среди щупалец актиний и других коралловых полипов. В этих случаях рачки оказываются защищенными от врагов стрекательными клетками щупалец кишечнополостных. Маленькие (самки до 5—6 мм, самцы обычно до 1,2 мм) крабики из семейства Hapalocarcinidae образуют на стволах рифообразующих кораллов известковые галлы, в полости которых обитают парами. Молодой крабик выедает тело полипа из его известковой чашечки и занимает его место. Растущая колония постепенно замуровывает крабика, но остается отверстие, через которое в полость галла попадает вода, а вместе с ней кислород для дыхания и органическая взвесь для питания. Среди игл морских ежей укрываются другие мелкие крабики, в некоторых случаях объедающие мускулы, приводящие в движение иглы и педицел-лярии ежа. Ряд креветок из подсемейства Ponton iinae, остальные представители которого также связаны с другими животными (губками, обо-л очниками, кишечнополостными), приспособлен к жизни в чашечках морских лилий, под цвет которых креветки окрашены. Среди крабов такими же обычными сожителями других животных можно считать так называемых «крабовнгороши-нок»(сем. Pinnotheridae), характеризующихся малыми размерами и относительно очень широким карапаксом. Различные виды этого семейства обитают внутри самых разных животных — в задней кишке морских ежей и голотурий, в мантийной полости двустворчатых моллюсков,; в асци-диях, а иногда в трубках полихет и в норах различных червей и раков.
Десятиногие ракообразные использует других животных не только в качестве убежища, но и для маскировки оружия, направляемого против своих врагов. Мелководные крабы из семейства Dorippi-dae при помощи загнутых на спинную сторону грудных ножек последней пары таскают над собой створку или обломок раковины какого-нибудь двустворчатого моллюска, полностью закрывая свое тело, за что и получили меткое название «стыдливых крабов». Крабы из семейства Majidae срезают своими клешнями различных обрастающих камни животных — губок, гидроидов, мшанок, колониальных асцидий — и помещают их на спинную сторону карапакса, где животные продолжают расти. Этому способствуют специальные крючковидные щетинки поверхности карапакса, к которым хорошо прикрепляется живой наряд краба, а также секрет желез задних ногочелюстей, склеивающий в кучки оторванных клешнями животных.
Сожительство раков-отшельников с актиниями — обычное явление. Для раков-отшельников актинии служат средством защиты от BparoBj актинии питаются остатками пищи своих хозяев. Поскольку как раки-отшельники, так и актинии получают от совместной жизни обоюдную пользу, это сожительство считается ярким примером одного из типов симбиоза — мутуализма (см. том 1).
Некоторые виды раков-отшельников и актиний могут жить как вместе, так и порознь. В таких случаях раки-отшельники при смене раковины оставляют свою сожительницу на старой брошенной раковине. Другие виды раков-отшельников (Pagurus arrosor) иногда существуют без актиний, но если рак находит актинию, то немедленно сажает ее на свою раковину, а при смене жилища пересаживает на новую. Наконец, рак-отшельник Pagurus prideauxi и актиния Adamsia palliata всегда живут только совместно. Лишь молодые актинии сидят на камнях, а взрослые — только на раковинах раков-отшельников, иногда по нескольку экземпляров на одной раковине. После поселения на раке-отшельнике актинии ее подошва разрастается, покрывает почти всю раковину и часто даже продолжается за устье. Если на раковине находятся две или несколько актиний, они располагаются так, чтобы уравновесить друг друга и не свалить на бок раковину и ее обитателя. Меняя раковину, рак-отшельник осторожно и заботливо клешнями переносит на новую раковину свою сожительницу. Искусственно разделенные рак-отшельник и актиния порознь потребляют меньше кислорода, чем когда они живут совместно. Рак-отшельник постоянно поедает пищу, обстрелянную стрекательными клетками актинии. В связи с этим он приобретает иммунитет по отношению к ее яду. Впрыскивание этого яда переносится раком-отшельником безболезненно, в то время как такая же доза вызывает гибель краба Carci-nus.
Третьим сожителем рака-отшельника и актинии часто бывают многощетинковые черви. Nerei-lepas fucata обитает в раковине, занятой Pagurus prideauxi, a Nereis cyclurus — в раковинахf занятых P. ochotensis и P. armatus. Эти черви иногда встречаются и в других условиях—в норах различных животных и в пустых раковинах. Замечательно, что рак-отшельник не трогает «своего» червя, хотя охотно поедает других. Больше того, при переселении в новую раковину он переносит с собой и червя. Черви принимают участие в трапезах рака-отшельника, высовываясь в это время из раковины и подбирая куски разрываемой хозяином пищи. Они приносят пользу своему сожителю,; очищая полость его раковины и объедая паразитов с его мягкого брюшка.
Раки-отшельники сожительствуют не только с актиниями, но и с представителями другой близкой группы кишечнополостных — зоантариями, играющими в этом симбиозе такую же роль, как актинии. Некоторые виды актиний и зоантарий живут только на определенных видах раков-от-шельников, другие, как например Sagartia parasitica* сожительствуют с вшогими видами.
Повшмо кишечнополостных, раковины раков-отшельников часто обрастают губки. При этом очень часто губка почти полностью растворяет раковину, и рак-отшельник оказывается окруженным губкой, как, например, у дальневосточного Pagurus cavimanus.
Защитное значение имеет, по-видимому, и способность некоторых десятиногих издавать звуки. В этом отношении особенно интересны раки-щелкуны (сем. Alpheidae), широко распространенные на тропических и субтропических мелководьях и встречающиеся также у нас в Черном и Японском морях. Они обычно прячутся между стволами кораллов, среди колоний мшанок, водорослей и камней, а также в губках. Одна из клешней этих рачков очень крупная и имеет своеобразное строение. На внутренней поверхности неподвижного пальца расположено углубление, а на подвижном пальце — соответствующий ему выступ. Перед щелчком подвижный палец отгибается, а затем благодаря сокращению сильного мускула прижимается к неподвижному. Выступ подвижного пальца входит в углубление неподвижного, и через канал, возникающий при смыкании пальцев, выбрызгивается струя выдавливаемой из углубления воды. При этом раздается громкий щелчок, усиливающийся благодаря пустым пространствам внутри клешни, которые служат резонаторами. Поскольку альфеу-сы обычно образуют плотные скопления г одновременное щелканье сотен рачков создает сильный шум. По описанию Н. И. Тарасова, «при извлечении губки, густо населенной альфеусами, слышна долго неутихающая громкая трескотня, напоминающая горение хвороста»4 а в других случаях при щелканье альфеусов «возникают шумы, напоминающие звуки, которые раздаются при заклепывании котла».
При щелчке возникают не только звуковые, но и ультразвуковые колебания. По всей вероятное-ти2 биологический смысл щелканья заключается в том, что мелкие ракообразные и рыбы получают сильный^; а иногда и смертельный ультразвуковой удар. Таким образом, щелканье можно считать защитной реакцией: недаром альфеусы отвечают им на всякого рода раздражения.
Лангусты также надают громкие звуки, отпугивая ими своих врагов. На первом членике задних антенн у лангустов есть лопасть, которая скребет о килевидный выступ передней части карапакса, причем возникает трескучий звук. Калифорнийский лангуст перед стрекотаньем угрожающе машет задними антеннами, усаженными острыми шипами. Рыбы отскакивают от лангуста и, по наблюдениям Моул1она, иногда теряют при этом свою добычу, которую подбирает лангуст. Известны и некоторые другие стрекочущие и скрипящие десятиногие.
К защитным приспособлениям десятиногих принадлежит также присущая многим из них способность к автотомии — произвольному отрыванию своих конечностей. Когда враг хватает за ногу какого-нибудь рака, Лт оставляет свою конечность врагу, а сам на оставшихся ногах спасается бегством. Привязанный эа ногу краб отрывает ее при виде осьминога. Таким обраэом можно лишить краба шести ног, но, если краба ослепить, автотомии не происходит. Отрывание конечностей осуществляется всегда в строго определенном месте, недалеко от их основания, при помощи специальных мышц — автотомизаторов, которые загибают ногу вверх до тех пор, пока она не обломится. В хместе отрыва внутри конечности находятся два расположенных поперечно соединительнотканных листка, помогающих быстро приостановить кровотечение. Некоторые десятиногие могут в случае необходимости отрывать себе конечности своими клешнями или задними ногочелюс-тями.
Способность к автотомии всегда сопровождается способностью к регенерации. Утраченная конечность постепенно отрастает вновь, тем скорее, чем чаще животное линяет, поскольку рост регенерирующей конечности происходит только после линьки. У крупных омаров полная регенерация клешней происходит через 2 года после их потери, у самок речных раков — через 3—4 года, а у чаще линяющих самцов — через 1V2—2 года. Если при утере конечности или глазного стебелька были повреждены связанные с ними нервные центры, то на их месте может вырасти придаток ненормального строения. Так, у многих креветок взамен глазного стебелька появляется успк, напоминающий переднюю антенву. Такое явление яааывают гетероморфозом.
Почти все десятиногие ракообразные раздельнополы, причем самцы внешне хорошо отличаются от самок. У креветок* как правило, самцы мельче самок, а у ползающих десятиногих наоборот, и только у крабов, живущих в других животных (Hapalocarcinidae, Pinnotheridae), самцы, так же как у креветок, уступают по размерам самкам. Часто самцы имеют более сильно развитые клешни, помогающие удерживать самку при спаривании. Иногда, как, например, у краба-плавунца Portunus, самец и самка различаются по цвету, а у травяного краба Carcinus по форме карапакса.
Особенно четко проявляется половой диморфизм в строении брюшка и его придатков. Как уже указывалось, передние брюшные ножки самцов целиком или частично преобразованы в органы, при помощи которых самец переносит сперма-тофоры, прикрепляя их к половым отверстиям самки, а у крабов вводя их в ее половые пути. Нередко брюшко самок бывает шире, чем у самцов (самки носят под ним икру), а у крабов оно, кроме того, часто состоит из большего числа сегментов, так как у самцов брюшные сегменты частично сливаются между собой.
У некоторых крабов наблюдается полиморфизм самцов. Так, среди самцов Inachus различаются три категории: мелкие крабики с толстыми клешнями, несколько более крупные, но с плоскими, как у самок, клешнями и самые крупные, опять с толстыми клешнями, причем семенники хорошо развиты только у крабов первой и третьей категории.
Гермафродитизм свойствен лишь немногим креветкам. Таковы все представители семейства Pandalidae, в том числе и промысловые виды Pandalus borealis и P. latirostris. Это типичные протерандрические гермафродиты. При наступлении половозрелости они становятся самцами, а на третий год жизни превращаются в самок. Сходным образом происходит смена пола у Lysmata seticau-da и Calocaris macandrae.
Самцы водных десятиногих находят самок, руководствуясь чувством обоняния и осязания. На жгутах передних антенн самцов креветок особенно много чувствительных щетинок, помогающих, вероятно, в поисках самок. Сухопутные крабы пользуются для этого зрением. Самцы издали видят самок и быстро бегут к ним.
Непосредственно перед спариванием самка линяет. У многих крабовидных отшельников, например у камчатского краба и у некоторых настоящих крабов, в том числе у мохнаторукого краба, линьке самки предшествует так называемое «положение рукопожатия», при котором самец и самка стоят друг перед другом и самец крепко держит в своих клешнях клешни самки. При этом оба партнера приподнимаются на своих грудных ногах. «Положение рукопожатия» может длиться довольно долго, у камчатского краба от 3 до 7 дней, и заканчивается линькой самки, причем, по некоторым наблюдениям, самец помогает самке слинять, стаскивая с нее старый панцирь. Другие крабы ждут линьки самки; взобравшись на нее сверху.
После линьки самки наступает спаривание. Самец при помощи передних брюшных ног прикрепляет сперматофоры близ половых отверстий самки и только у крабов вводит их в семяприемники самки. Самцы некоторых раков-отшельников приклеивают сперматофоры к раковине^ в которой живет самка.
У разных видов спаривание осуществляется по-разному. Креветки Sicyonia carinata и Athanas nitescens располагаются перпендикулярно друг к Другу, причем самец оказывается лежащим на спине снизу, а самка спиной кверху над ним. У речных раков, омаров, лангустов, Galatheidea и некоторых других самка лежит на спине, а самец находится над ней спиной вверх. У многих крабов и у крабовидных отшельников самец сидит сверху, на спинной стороне самки. Молодые самцы (длиной 10 мм) Athanas nitescens в течение нескольких минут спариваются с разными самками 4—5 раз, более крупные (17—20 мм) — один раз. Самцы других креветок, китайского и камчатского крабов, речного рака Cambarus также часто спариваются с несколькими самками. Самкам некоторых крабов для оплодотворения яиц достаточно одного спаривания в несколько лет,, так как сперматозоиды в семяприемниках остаются живыми. Самка американского голубого краба (Callinectes) спаривается один раз в жизни, в возрасте 3 лет2 а живет 7 лет и откладывает яйца ежегодно.
Промежуток между спариванием и откладкой япц у разных видов различен. Самки Athanas и других креветок начинают откладку яиц через несколько минут после спаривания, речные раки— через 10—45 суток, краб Carcinus — через несколько месяцев. Для откладки яиц омары, речные раки и раки-отшельники опрокидываются на спину, а креветки ложатся па бок и подгибают брюшко так, чтобы между ним и нижней поверхностью груди образовалось пространство, куда и попадают яйца. Вместе с яйцами выделяется секрет, растворяющий оболочки сперматофоров, сперматозоиды освобождаются и оплодотворяют яйца. Самки лангустов разрывают оболочку сперматофора когтем задней ходильной ноги. Секрет цементных желез, находящихся у крабов в семяприемниках, а у остальных десятиногих на нижней поверхности брюшка и на брюшных ножках, образует наружную оболочку каждого яйца, а также тяж, который прикрепляет его к брюшным ножкам матери. Крабы Carcinus при откладывании яиц зарываются в грунт4 без чего не происходит прикрепления яиц. В дальнейшем яйца останутся на плеоподах вплоть до выхода личинок или молодых рачков. Только у самых примитивных креветок, объединяемых в семейство Penaeidae, процесс откладывания яиц происходит иначе: они откладывают яйца прямо в воду.
Плодовитость десятиногих колеблется в широких пределах. При этом чем больше яиц, тем меньше их размеры. Очень плодовиты некоторые крабы: число яиц, выметываемых Cancer pagurus Carcinus maenasj достигает 3 млн. (но в Черном море у последнего не более 180 ООО яиц), у Calli-nectes sapid us — 2 млн.А у Eriocheir sinensis оно колеблется от 270 ООО до 920 ООО и т. д. Гораздо ниже плодовитость наших северных крабов (Hyas coarctatus — 1 000—16 000 яиц, Н. araneus — до 110 000 яиц). Очень высока плодовитость лангустов — от 500 000 до 1,5 млн. яиц. У раков-отшельников число яиц обычно не доходит до 10 000, но у близкого к ним камчатского краба оно значительно больше — от 70 000 до 270 000. Среди креветок наиболее плодовиты Penaeidae, откладывающие яйца в воду: Penaeus trisulcatus мечет около 800 000, Parapenaeopsis longirostris около 400 000 яиц. Креветки, вынашивающие яйца на себе, откладывают их в меньшем количестве: Pandalus latirostris — 150—450, P. borealis — 400—3 000, Palaemon adspersus — 160—3600, P. elegans — 280—1600, Crangon crangon — 1500— 15 000. Сравнительно невелика и плодовитость европейского и норвежского омаров (8 000— 32 000 и 1 300—4 000 яиц).
У всех пресноводных и глубоководных десятиногих плодовитость значительно меньше, чем у морских мелководных. Наши речные раки откладывают от 60 до 500—600 яиц* тропические пресноводные крабы — 20—150* глубоководные планктонные креветки — 15—60л глубоководный Munidopsis rostrata — 230. tyfa их крупных яиц выходит не личинках а уже почти сформировавшийся рачок.
Яйца остаются прикрепленными к брюшным ножкам матери в течение продолжительного времени. Камчатский краб и европейский омар вынашивают их почти целый год, норвежский омар — 9 месяцев, краб Cancer pagurus — 7—8 месяцев, речные раки — около полугода, Pandalus borealis — 5 месяцев, Carcinus maenas — летом 2 месяца, зимой 5—6 месяцев, китайский мохнато-рукий краб и креветки рода Palaemon — 1—1V2 месяца, пальмовый вор — 3—4 недели, краб-плавунец Portunus trituberculatus — 2—3 недели. Эти различия зависят в первую очередь от температуры, которая влияет на скорость эмбрионального развития.
У большинства десятиногих из яйца выходит личинка, резко отличающаяся от взрослых как по строению, так и по образу жизни. У многих пресноводных, глубоководных и обитающих при низкой температуре видов развитие прямое, т. е. все личиночные стадии проходят под покровом яйцевой оболочки и из яйца появляется маленький почти сформировавшийся рачок.
Различают несколько стадий плавающих личинок десятиногих ракообразных. Первая из них — науплиус — свойственна только примитивному семейству креветок Penaeidae. Стадия метанауплиуса есть также только у них и у некоторых представителей близкого семейства Sergestidae. У большинства морских мелководных десятиногих из яйца выходит личинка, находящаяся на стадии зоэа. Для нее характерно разделение тела на покрытую карапаксом головогрудь и сегментированное брюшко, причем последний его сегмент еще не отделен от тельсона. Глаза зоэа стебельчатые, очень сильно развиты ногочелюсти, наружные ветви которых служат основными органами плавания, а задние грудные и все брюшные ножки еще не образовались или зачаточны. У зоэа крабов передняя часть тела вздутая, на карапаксе расположены длинные шипы — один на спине и два по бокам, а крометого, имеется длинный рострум. У зоэа раков-отшельников и близких к ним групп задний край карапакса выемчатый, его задне-боковые углы оттянуты в шипы и также есть длинный рострум. Все эти выросты помогают личинкам существовать в толще воды, так как затрудняют их погружение.
За стадией зоэа у длиннохвостых десятиногих следует стадия м и з и с. На этой стадии все грудные конечности хорошо развиты, снабжены наружными ветвями и служат для плавания, задний брюшной сегмент отделен от тельсона, начинается развитие брюшных конечностей. У большинства креветок стадия мизис сходна с мизидами, почему она и получила свое название, но у некоторых групп она имеет причудливую форму. Таковы, например, листовидные, совершенно прозрачные мизисные личинки лангустов, так называемые филлозомы, или шаровидные личинки глубоководных Егуо-nidae, так называемые эрионеикусы. У омаров личинка выходит из яйца на этой стадии. Мизисной стадии длиннохвостых соответствует стадия метазоэа крабов, раков-отшельников и близких к ним групп. Метазоэа отличается от мизис отсутствием наружных ветвей грудных ног.
Последняя личиночная стадия носит название декаподитной. На этой стадии кара-пакс уже почти такой же, как у изрослого ракообразного, но брюшко часто имеет иное строение. Декаподитная стадия крабов — мегалопа и декаподитная стадия раков-отшельников и близких к ним форм — глаукотоэ характеризуются еще не подогнутым под грудь и вполне симметричным брюшком, снабженным сильными плеоподами, при помощи которых личинка плавает. Затем она садится на дно, линяет и превращается в молодую особь с нормально устроенным брюшком, подогнутым под грудь у крабов или спирально закручепным у раков-отшельников.
Продолжительность существования личинок в планктоне различна у разных видов: у креветок Palaemon adspersus и Crangon crangon и краба Carcinus maenas — 4—5 недель, у пресноводной креветки Atyaephyra desmaresti и у европейского омара — 2—3 недели, у креветки Panda-lina brevirostris — 2 месяцаt у обыкновенного лангуста Palinurus — 3 месяца, у креветки Ser-gestes arcticus — 4—5 месяцев, а у лангуста Ра-nulirus — до 6 месяцев. За время пребывания в планктоне филлозомы лангустов разносятся течениями на далекие расстояния. С этим связывают циркумтропическое распространение некоторых лангустов.
Многие планктонные личинки десятиногих — хорошие пловцы. Когда они плавают при помощи ногочелюстей, т. е. на стадии зоэа, они двигаются задним концом вперед или, как многие зоэа крабов, спинной стороной вперед. При этом зоэа Galathea проплывает 1 м за 45—56 с, Porcellana — за 65—92 с. После превращения в последующие стадии, когда органами движения становятся плеоподы, личинки двигаются передним концом вперед. Пищей личинкам служат разные планктонные водоросли и животные.
Огромное количество личинок погибает, становясь жертвами различных хищников или попадая в неподходящие условия. По расчетам японского зоолога Марукава, из 200 000 зоэа первой стадии, которых дает одна самка камчатского краба, до второй стадии зоэа доживает 13,: а до стадии глаукотоэ 7 личинок.
У большинства пресноводных, глубоководных и холодноводных десятиногих яйца крупные, богатые желтком, и всё превращение личинки проходят под покровом яйцевой оболочки. Из яйца выходит маленький рачок, который отличается от взрослых главным образом отсутствием последней пары брюшных конечностей. Новорожденные рачки удерживаются на плеоподах матери при помощи своих конечностей. У речных раков северного полушария (сем. Astacidae) для этого служат клешни, а у речных раков южного полушария (сем. Parastacidae) — задние грудные ножки. Сходным образом ведут себя вылупившиеся из яиц пресноводные крабы (сем. Potamonidae), полуглубоководная креветка Sclerocrangon ferox и глубоководная S. zenkevitchi.
С размножением связаны миграции многих десятиногих. Они стремятся к моменту выхода из яиц личинок попасть в условия, наиболее благоприятные для существования своих новорожденных потомков. Мохнаторукий краб, поднимающийся довольно высоко вверх по рекам Западной Европы, для размножения переселяется в солоноватую воду предустьевых пространств. Точно так же мигрируют в море для размножения некоторые Penaeidae, остальное время проводящие в солоноватой или пресной воде, как например Penaues carinatus и Penaeopsis dobsoni из индийского солоноватоводного озера Чилька или Ре-naeus plebejus из рек Австралии. Многие морские креветки и некоторые крабы, когда приходит время выхода личинок из яиц, перекочевывают прибрежные мелководья, где вода сильнее прогрета. Наиболее интересны нерестовые миграции сухопутных десятиногих. Самки сухопутных раков-отшельников — пальмового вора и Соепо-bita, а также сухопутных крабов из семейства Gecarcinidae, несущие на брюшных ножках яйца, в которых сформировались зоэа, массами стремятся к морю. Они идут иногда издалека, преодолевая по нескольку километров. Переселения обычно совершаются ночью или днем во время дождя, поскольку период размножения сухопутных десятиногих приходится на сезон тропических ливней. Достигнув морского берега, самки заходят в воду или забираются на камни и, как пальмовый вор, стряхивают яйца в воду либо, частично погрузившись в воду, ждут, когда нз яиц вылупятся личинки. В дальнейшем личпнки развиваются в море, там оседают на дно и только через некоторое время выбираются на сушу.
Развитие личинок пальмового вора дает довольно полное представление об истории формирования этого вида. Планктонные зоэа и глаукотоз пальмового вора такие дее9 как у раков-отшельников. После оседания на дно, т. е. через 4—6 месяцев после вылупления из яйца, личинка пальмового вора имеет спирально изогнутое мягкое брюшко, которое она прячет в пустую раковину какого-нибудь морского брюхоногого моллюска. В это время она совершенно сходна с обычным раком-отшельником. Затем она перебирается на сушу и использует раковины наземных моллюсков, напоминая взрослые экземпляры близкого сухопутного рода Coenobita. Наконец, после очередной линьки, она оставляет раковину, в результате нескольких следующих линек ее брюшко постепенно укорачивается и подгибается под грудь, и таким образом она превращается из рака-отшельника в пальмового вора. Вероятно, так же, но в течение очень длительного времени шла эволюция этого вида и Coenobita, происшедших от морских раков-отшельников.
Закончившие превращение молодые десятиногие ведут такое же существование, как взрослые, с каждой линькой увеличиваясь в размерах. Сначала они линяют часто, потов^ промежутки между линьками удлиняются, и крупные особи таких видов, как омар и камчатский краб, линяют один раз в два года. Линька контролируется органами внутренней секреции, находящимися в глазных стебельках. Удаление глазных стебельков вызывает более частые линьки, так как в кровь перестает поступать тормозящий их гормон.
Перед линькой под старым панцирем формируется новый, еще мягкий, не содержащий солей кальция. Старый панцирь в процессе линьки лопается на границе между грудью и брюшком, а у крабов, кроме того, по бокам грудного отдела. Длиннохвостые десятиногие вытаскивают через образовавшуюся щель сначала переднюю, а затем заднюю части тела, крабы сначала брюшко, потом голову и грудь, ходильные ноги и, наконец, клешни. Речные раки при этом, как впервые описал еще Реомюр в 1712 г., опрокидываются на спину и начинают резко сгибать брюшко и конечности, вследствие чего и обра зуетси щель в старом панцире. Возможно, что при вытаскивании из старого панциря тела и конечностей они механически вытягиваются и рост осуществляется непосредственно в процессе линьки. Она занимает обычно 10—30 мин, но затвердевание нового панциря требует гораздо более продолжительного времени: у камчатского краба 10 дней, у омара 20—50 дней, у мохнато-рукого краба больше месяца, у краба Cancer pagurus 2—3 месяца. При этом соли кальция поступают из крови в новый панцирь и пропитывают его. Перед линькой они накапливаются либо в печени (у большинства десятиногих), либо в так называемых г а с т р о л и т а х — двух овальных твердых образованиях* находящихся в желудке речных раков.
До тех пор пока не ватвердел новый панцирь, десятиногие остаются совершенно бевзащитными. Кроме того, они не могут питаться, так как все их ротовые придатки и перетирающий пищу аппарат желудка еще мягкие. Многие десятиногие на это время забиваются под камни или в другие убежища, где они оказываются в безопасности, и там мужественно переживают тяжелый для них период жизни.
Некоторые десятиногие живут очень долго: американский омар до 50 лет, широкопалый речной рак до 20 лет, камчатский краб до 23 лет* лангуст более 15 лет, мохнаторукий краб до 6,, иногда до 10 лет. Пальмовый вор длиной более 10 см имеет от роду не менее 5 лет. Продолжительность жизни креветок обычно не превышает 4 лет (Crangon crangon, Pandalus), а мелкие виды не доживают и до года.
Подавляющее большинство десятиногих обитает в области морского мелководья. Особенно богата ими фауна тропиков. Коралловые рифы населены бесчисленным множеством самых разнообразных крабов, креветок и раков-отшельников, часто очень пестро окрашенных и имеющих причудливую форму. По направлению на север и на юг от тропической 8оны фауна десятиногих беднеет. В пределах умеренных широт по богатству десятиногими выделяется северная часть Тихого океана, где обитает наибольшее число родов и видов таких семейств креветок, как Сгап-gonidae, Hippolytidae, Pandalidae, а также кра-бовидных отшельников Lithodidae. В Беринговом море известно 117, в Охотском 113, в Японском 82 вида десятиногих. Из северной части Тихого океана шло расселение некоторых групп десятиногих в Полярной бассейн и даже в северную часть Атлантического океана. Очень бедна десятиногими ракообразными Антарктика, откуда указано всего 8 видов.
Морские десятиногие обитают от приливо-отливной полосы до глубины 5300 м. В этом отношении они уступают многим другим отрядам ракообразных (бокоплавам, равноногим и др.), которые приспособились к жизни и на больших глубинах, вплоть до максимальных (11 км). Среди глубоководных донных десятиногих много древних родов, отсутствующих на мелководьях, но известных в ископаемом состоянии (Eryonidae, крабы Prosoponidae и Ilomolidae и др.). Очень характерны глубоководные планктонные креветки, принадлежащие к семействам Oplopho-ridae, Penaeidae, Sergestidae и др. Обычно они темно-красного цвета, их конечности удлинены, и многие из них обладают способностью к свечению.
У многих глубоководных Sergeatidae на конеч-ностях* вдоль боковых краев карапакса и на брюшной стороне тела располагается до 150 фо-тофоров. У некоторых видов часть фотофоров снабжена линзами. Среди глубоководных креветок известны виды, лишенные фотофоров, но тем не менее светящиеся. Их покровы очень тонкие, и сквозь них просвечивают участки печени, превратившиеся в органы свечения — так называемые органы Песта. Наконец, некоторые креветки, преимущественно из рода Systellaspis, выбрасывают струю светящейся жидкости и при помощи «огневой завесы» скрываются от врагов. Присутствие фотофоров, расположение и число которых для каждого вида строго постоянно, помогает в полном мраке океанических глубин самцам находить самок и всем вместе собираться в стаи.
Приливо-отливная полоса тропических морей населена очень своеобразными десятиногими4 из которых наиболее характерны манящие крабы. Это мелкие (ширина карапакса 0,8—За5 см) кра-бики* живущие на илистом грунте большими колониями и замечательные тем, что их самцы своей огромной клешней или обеими клешнями совершают сложные манящие движения, ритмически разводя клешни в стороны, поднимая и опуская их и располагая их перед собой. Манящие движения в деталях различны у разных видов. У видов рода Uca с узким лобным краем преобладают движения клешней в вертикальной, у видов с широким лобным краем — в горизонтальной плоскости. Приближение самки или другого самца вызывает более энергичные манящие движения. Изучение их у разных видов (с применением киносъемки н иных современных методов) указывает на многообразие биологического значения этих движений. Во-первых, таким образом самцы отпугивают других самцов, сообщая им о том, что данный участок уже занят. Во-вторых, в период спаривания манящие движения самца служат для привлечения самки. В промежутках между этими движениями самец постепенно приближается к самке, которая сначала принимает позу защиты и отступает, а затем сама начинает двигаться навстречу самцу. Прп этом оба партнера размыкают н смыкают пальцы своих клешней. Если какой-нибудь самец не обратит внимания на предупреждение и вторгнется на чужую территорию, между ее хозяином и пришельцем возникает ожесточенное сражение. Если перед самцом поставить зеркало, он бросается в бой со своим отражением, что указывает на ведущую роль зрения во всех этих явлениях. У манящих крабов рода Dotilla победитель после окончания драки исполняет триумфальный танец. Нередко драки между двумя самцами происходят из-за самки, которая в таких случаях сама удаляется и ждет исхода боя. Замечательно, что совершенно сходные повадки свойственны некоторым представителям другого семейства крабов — Grapsidae^ живущим также в приливо-отливной иолосе тропических морей. Сравнительное исследование манящих движений приводит к выводу о том, что они развились независимо у представителей двух разных семейств из поз защиты и нападения, причем впоследствии к первоначально исключительно агрессивному их назначению добавилось сексуальное.
Некоторые морские десятиногие могут переносить значительное опреснение; среди них есть виды, обитающие преимущественно в опресненных участках моря и способные заходить в совершенно пресную воду. Таковы, например, мохнаторукие крабы рода Eriocheir. Один из них — Е. japonicus — живет перед устьями рек от Амурского лимана до Гонконга и Тайваня, а также Северо-Восточного Сахалина, другой — Е. sinensis — в таких же условиях в Китае. Китайский мохна-торукий краб в балластных цистернах судов был случайно перевезен около 1912 г. в бассейн Эльбы. Оттуда он начал быстро расселяться вдоль западных берегов Европы и сейчас многочислен во многих реках и их устьевых частях в ФРГ, ГДР, Бельгии, Нидерландах, Франции, Даниих Швеции и Финляндии; оп разрушает своими норами плотины, рвет рыболовные сети и портит пойманную в них рыбу. Другой солоноватовод-нбгй крабик — Rhithropanopeus harrisi — таким же образом попал из предустьевых пространств американских рек в Нидерланды, а затем проник в Балтийское, Азовское и Каспийское моря, где силыю размножился и используется в пищу рыбами.
В пресных водах обитает сравнительно немного видов десятиногих. Среди них широко известны речные раки, принадлежащие к трем семействам — Astacidae, Parastacidae и Austrastacidae. Astacidae живут в умеренной и отчасти южной полосе северного полушария, Parastacidae — в умеренной полосе южного полушария — в Южной Америке, Австралии, а также на острове Мадагаскар, Austrastacidae — только в южной части Австралии. Почти па всем протяжении тропиков речных раков нет. В пределах СССР речные раки представлены двумя родами — Astacus и Cambaroides. Род Astacus занимает почти всю Европу и западную часть Азпп, род Cambaroides — бассейн Амура, северную Японию1 Корею и Сахалин.
Наиболее широко распространены и имеют довольно важное промысловое значение два наших вида речных раков — широкопалый (А. astacus) и узкопалый (A. leptodactylus). Широкопалый рак живет главным образом в водоемах, относящихся к бассейну Балтийского моря; он обычен в Латвии, Литве, Эстонии, в Ленинградской области, в некоторых районах Украины и Белоруссии. Область распространения узкопалого рака гораздо шире и охватывает водоемы бассейнов Каспийского, Черного и Азовского морей, Каспийское и Азовское, моря, а также реки и озера Западной Сибири.
Шпрокопалый и узкопалый раки совместно не встречаются, и их ареалы не перекрывают друг друга. В прошлом веке узкопалый рак начал расселяться вверх по бассейну Волги, вытесняя широкопалого. Известны многочисленные случаи посадок раколовами узкопалого рака в озера, населенные широкопалым. Проходит 10—20 лет. и широкопалый рак исчезает^ полностью замещаясь узкопалым.
Оба вида сходны по своей биологии. Они живут в воде со сравнительно высоким содержанием кислорода и минеральных солей, питаются преимущественно растительной пищей, имеющейся обычно в избытке. Узкопалый рак несколько более плодовит и более вынослив по отношению к химизму воды и ее кислородному режиму. Однако это еще не объясняет, каким образом он неизменно вытесняет широкопалого. Не удалось получить ответ на этот вопрос и при оовместном содержании обоих видов в бассейнах. Время от времени раки дрались, но драки между особями разных видов происходили не чащеЛ чем менаду особями одного вида.
В Литве, где встречаются и широкопалый и узкопалый раки, удалось заметить, что на стыке их ареалов обитают особи, по своим признакам занимающие промежуточное положение между обоими видами. Я. М. Цукерзис считает их гибридами и допускает возможность постепенного исчезновения признаков того или иного вида у потомков этих гибридов. В искусственных условиях удалось скрестить между собой оба вида и получить гибридную молодь, но она не доросла до взрослого состояния. Следует при этом иметь в виду, что узкопалый и широкопалый раки различаются не только по форме клешней, ростру-ма, вооружению карапакса и т. д., но и по числу хромосом: у узкопалого рака их 184, а у широко-палого около 100. Для растений давно известно, что полиплоиды характеризуются повышенной жизнеспособностью, плодовитостью и другими преимуществами; по-видимому, это подтверждается и на примере речных раков.
На численность и распространение речных раков большое влияние оказали их болезни^ в первую очередь, рачья чума. В 1870 г. она появилась во Франции, затем продвинулась восточнее, а в 1890 г. проникла в западную часть России. К 1898 г. эпизоотия рачьей чумы захватила всю Россию, включая Западную Сибирь. Во многих местах раки вымерли поголовно и исчезли, в других их запасы постепенно начали восстанавливаться. Но в 1919—1920 гг. возникла новая вспышка рачьей чумы, опять уничтожившая огромное количество раков.
Заболевшие животные ходят на выпрямленных ногах, выползают из нор днем, чего здоровые раки никогда не делают, через некоторое время у них начинаются судороги, и в течение недели ракп погибают. Возбудителем болезни служит грибок Aphanomyces astaci, развивающийся в покровах и в центральной нервной системе больных раков. Болезнь распространяется посредством ооспор этого грибка.
В пресных водах тропиков взамен отсутствующих там речных раков водится много различных крабов и креветок. К пресным водам целиком приурочено богатое видами семейство крабов Potamo-nidae, включающее около 20 родов и более 200 видов. Некоторые виды этого семейства встречаются даже в высокогорных водоемах (до 2100 м над уровнем моря в Индии). В Советском Союзе обитает 11 видов этого семейства. Они обычны у нас в Крьшу, на Кавказе и в Средней Азии. В сырых лесах Закавказья они встречаются во влажной земле, в которой роют норы. Среди креветок есть также одно целиком пресноводное семейство — Atyidae, характеризующееся присутствием пучков щетинок на пальцах клешней, при помощи которых рачки захватывают грунт. Географическое распространение этого семействаг а также его морфологические особенности указывают на его глубокую геологическую древность. В бассейне Амура водится два вида атиид, представляющих вместе со многими рыбами и моллюсками тропический элемент в амурской фауне. Еще одна креветка из этого семейства — Troglocaris anoph-thalmus — живет в пещерных водоемах Закавказья. Кроме этих двух семейств, в тропических пресных водах обитают виды крабов и креветок, относящиеся к преимущественно морским родам.
Особый интерес представляют пещерные десятиногие. Известно около 20 слепых пещерных видов Atyidae и 10 видов Palaemonidaej разбросанных по различным континентам и островам. В пещерах США живет 18 видов слепых речных раков, а в Мексике, на Новой Гвинее и острове Калимантан — пещерные крабы семейств Pota-monidae и Gecarcinidae. Наиболее удивительным следует считать существование представителя рода Munidopsis (сем. Galatheidae) в солоноватом подземном озере на одном из Канарских островов; все остальные многочисленные виды этого рода обитают на больших океанических глубинах. Очевидно, и они и подземный М. polymorpha некогда произошли от вымерших мелководных морских предков и дожили до наших дней в подземных и глубинных водах, как в убежищах, сохранивших много различных остатков древних фаун. Другой загадкой остается разорванный ареал примитивных креветок рода Procaris, обитающих в лужах с морской водой: один вид найден на острове Вознесения в Атлантическом океане, другой — на одном из островов Гавайского архипелага в Тихом океане.
Десятиногие используются в пищу людьми и интенсивно добываются почти во всех странах. В СССР особенно большое экономическое значение имеет промысел камчатского краба (Рага-lithodes camtschatica), из мускулов ног которого делают консервы. Он принадлежит к семейству Lithodidae, т. е. крабовпдных раков-отшельников, и населяет Японское, Охотское и Берингово моря.
Ширина карапакса самцов этого вида в среднем 16 см« а у некоторых экземпляров достигает 25 см. Расстояние между концами средних ходильных ног таких крупных особей — 1V2 м, а масса их тела — 7 кг. Самки значительно мельче (ширина карапакса до 16 см), и их использование для изготовления консервов нецелесообразно.
Всю свою долгую жизнь камчатские крабы проводят в странствованиях, причем каждый год повторяют один и тот же маршрут. Их миграции в значительной степени зависят от колебаний температуры. Для самцов наиболее благоприятна температура между 2 и 7 °С, хотя они выдерживают некоторое время и колебания от —2 до +18 °С. Оптимальная температура для самок несколько выше. Если температура повышается или понижается за пределы оптимальной, крабы перемещаются в районы, более подходящие для них по тепловому режиму. Другой причиной миграций служит размещение кормовых животных: крабы кочуют в поисках пищи, пригодной для той или иной фазы их жизненного цикла.
Наибольшие скопления крабов сосредоточены у западной Камчатки, где в основном и ведется их промысел. Здесь крабы проводят зиму вдали от берегов, на значительной глубине 110—200 м, куда зимнее охлаждение еще не успевает распространиться и где господствует низкая положительная температура. Весной начинается прогрев воды на мелководьях, а охлаждение достигает мест зимовки крабов. Это и заставляет крабов начинать переселение с больших глубин к берегам.
Скорость передвижения крабов невелика. За сутки они проходят по прямой не более 5—7 миль. При этом разные особи двигаются в разных направлениях, и все стадо в целом обычно перемещается за сутки всего на 1—2 мили. Каждое стадо кочует в довольно ограниченном районе, площадь которого составляет у Камчатки около 100, а в Приморье около 40 кв. миль. У охотоморскнх берегов Камчатки различаются 4 таких района и соответственно 4 стада, из которых наиболее многочисленно северное* приуроченное к Хайрюзов-скому району.
С мест зимовки к берегам крабы двигаются отдельными косяками — крупные самцы отдельно от самок и молоди. Самки несут на своих брюшных ножках яйца, содержащие уже вполне сформировавшихся личинок. По дороге на мелководья личинки выходят из яиц,, а самки продолжают свой путь.
Приблизительно через месяц после начала миграции косяки самцов и самок встречаются на мелководьях (5—30 м), где температура воды в это время достигает 2—4°С, и перемешиваются. Самцы располагаются перед самками и держатся своими клешнями за их клешни, принимая «положение рукопожатия». Оно длится от 3 до 7 суток. Затем самцы помогают самкам слинять, стаскивая с них загрязненный старый панцирь, после чего происходит спаривание. Самец отбрасывает старый панцирь и прикрепляет к основаниям третьей пары ходильных ног только что слинявшей самки сперматофор — лентовидную массу спермы, покрытую нерастворимой в воде оболочкой, после чего оба партнера расходятся.
Через несколько часов, а иногда и дней после спаривания самка откладывает на свои брюшныо ножки икру, которая при атом оплодотворяется и которую самка носит до следующей весны, т. е. приблизительно 111/2 месяцев.
После спаривания косяки самцов и самок опять мигрируют раздельно. Если весеннюю миграцию можно назвать нерестовой, то' летние должны считаться кормовыми. Перед ними самцы линяют, но, в отличие от самок, в одиночестве, среди каменистых россыпей, обычно против далеко выдающихся в море мысов.
Со второй половины июля и до конца сентября у берегов Камчатки и с конца мая—начала июня до ноября в заливе Петра Великого крабы совершают миграции, переходя с одного кормового поля на другое. Самцы держатся глубины 20—60 м и температуры 2—7 °С. Самки перемещаются несколько ближе к берегам. Осенью прибрежные воды начинают охлаждаться, и крабы отступают от берегов, пока не достигнут мест зимовки. Следующей весной все повторяется сначала. Вышедшая из яйца первая личинка — пр то-зоэа — линяет через несколько минут после вы-лупления и превращается в зоэа, которая живет в толще воды около 2 месяцев, линяет 4 раза и переходит в глаукотоэ. Глаукотоэ оседает на дно и в течение 20 дней ползает в зарослях водоросли анфельции, среди гидроидов и т. п. Глаукотоэ уже имеет все конечности взрослого краба.
После линьки она превращается в молодого краба — малька, вполне сходного со взрослыми, но имеющего относительно более длинные шипы на панцире. Ширина его тела 2 мм. Малек живет так же и в таких же условиях, как глаукотоэ, и несколько раз линяет. Только через 3 года он покидает заросли водорослей и гидроидов и переходит на участки песчаного грунта. К 6—7 годам, достигнув ширины панциря около 7 см, молодые крабы начинают собираться в косяки и мигрировать.
Самки растут несколько медленнее, чем самцы, но зато раньше достигают половозрелости. Уже на восьмом году жизни они начинают откладывать икру, а самцы становятся половозрелыми только в возрасте 10 лет. В это время они достигают ширины панциря 12,5 см и годятся для изготовления консервов.
Вследствие позднего наступления половозрелости и медленного роста камчатского краба его запасыг подорванные интенсивным промыслом, восстанавливаются с большим трудом. Из огромного количества ежегодно рождающихся личинок до взрослого состояния доживают очень немногие. Остальные становятся жертвами различных хищников или уносятся течениями. Заросли, подходящие для оседания личинок, есть только в Хап-рюзовском районе охотоморского побережья Камчатки. Отсюда молодь краба заселяет более южные участки. Однако в годы с пониженной температурой воды на мелководьях такая миграция невозможна, и южные стада камчатского краба совсем не получают пополнения.
В 30-е гг. было сделано предложение об акклиматизации камчатского краба в Баренцевом море. В настоящее время есть сторонники и противники этой идеи. Методика перевозки крабов и их личинок с Дальнего Востока разработана, перевозка осуществлена, и сейчас ведутся наблюдения в акварнальных и естественных условиях над поведением камчатского краба в новом для него водоеме.
Краба ловят ставными сетями, которые стремятся поставить так, чтобы перегородить основные пути миграции. Небольшие суда привозят сети с пойманными крабами на краболов, где производят выпутку крабов, отрывают у них ноги и клешни, варят их, рубят, вытряхивают мясо и изготовляют из него консервы. Помимо камчатского краба, в наших дальневосточных морях в меньшем количестве добывают креветок из рода Pandalus. В пресных водах, преимущественно на Украине, ведется промысловый лов речных раков.
За рубежом добывают много различных десятиногих, из которых на первом месте стоят креветки. В 1962 г. их было поймано 700 тыс. т, причем особенно развит этот промысел в Китае, США, Индии, Японии. Наибольшее значение в уловах имеют семейства Penaeidae, Pandalidae и Crangonidae. В некоторых странах Южной и Юго-Восточной Азии ведется искусственное разведение креветок* преимущественно пенеид. Прибрежные низменные пространства при помощи дамб превращают в цепь прудов, соединяющихся каналом с морем. В прилив вода заливает пруды и остается там благодаря тому,что шлюзы во время отлива закрывают. В такие пруды креветки заходят сами, или туда выпускают пойманных в море личинок. Креветок подкармливают рисовыми отрубями. При таком ведении хозяйства удается получать до 500 кг креветок с гектара.
Меньшее значение имеет промысел настоящих крабов, особенно развитый в США. Их продают свежезамороженными, вареными и в виде консервов, причем высоко ценятся недавно слинявшие особи. Из других десятиногих считаются деликатесом лангусты и омары. Запасы европейского омара подорваны промыслом, и Канада экспортирует в Европу довольно значительное количество американских омаров. Для восстановления запасов омара производится его разведение в специальных садках.
Наша страна использует десятиногих еще совершенно недостаточно. Нет сомнения в том, что в будущем их промысел у нас значительно возрастет.
Книги:
 Жизнь животных. Том 2. Моллюски. Иглокожие. Членистоногие / В. Е. Соколов. — 1988
Жизнь животных. Том 2. Моллюски. Иглокожие. Членистоногие / В. Е. Соколов. — 1988
 Зоология беспозвоночных. От артропод до иглокожих и хордовых. / Вестхайде В., Ригер Р. — 2008
Зоология беспозвоночных. От артропод до иглокожих и хордовых. / Вестхайде В., Ригер Р. — 2008



 Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008