Тип Caudovirales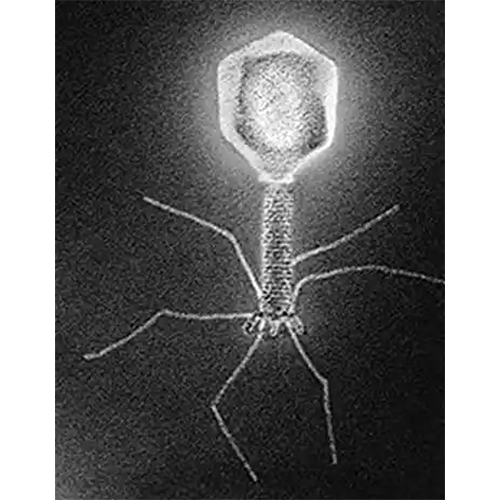 Caudovirales
Caudovirales
CaudoviralesТип Herpesvirales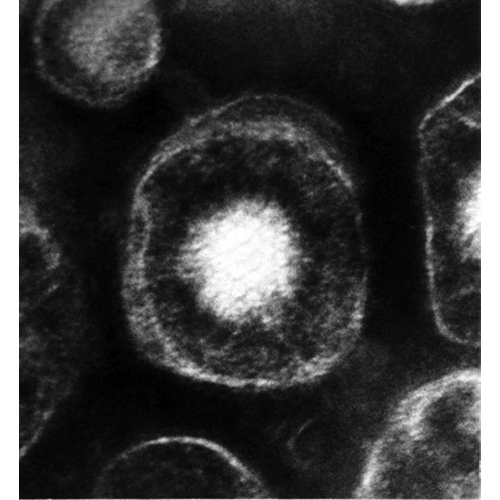 Herpesvirales
Herpesvirales
HerpesviralesТип Mononegavirales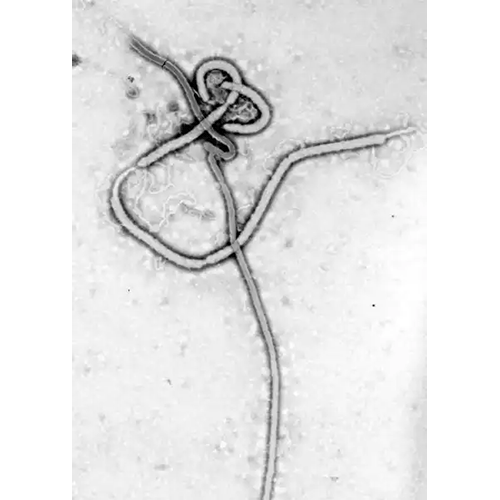 Mononegavirales
Mononegavirales
MononegaviralesТип Nidovirales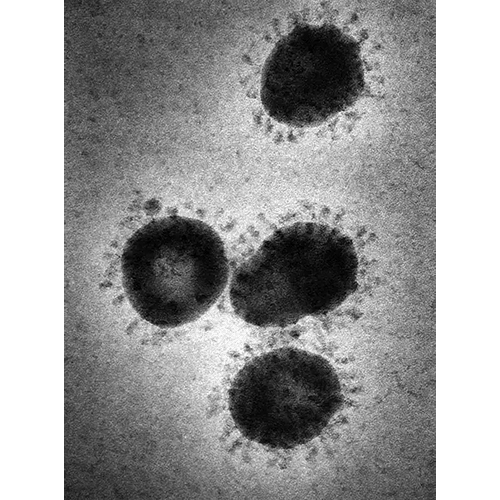 Nidovirales
Nidovirales
NidoviralesТип Picornavirales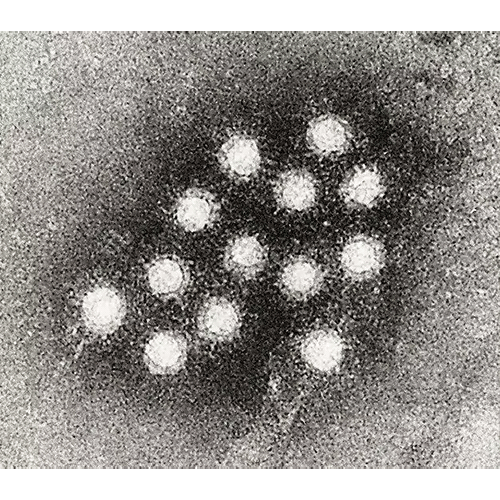 Picornavirales
Picornavirales
PicornaviralesТип Tymovirales Tymovirales
Tymovirales
TymoviralesТип НЕ ОПРЕДЕЛЕНО Not assigned
Not assigned
Not assigned