


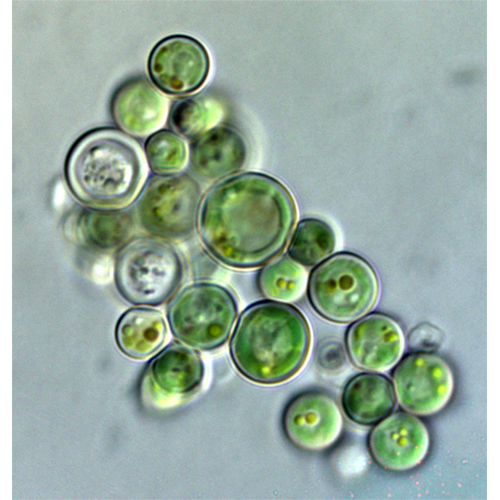

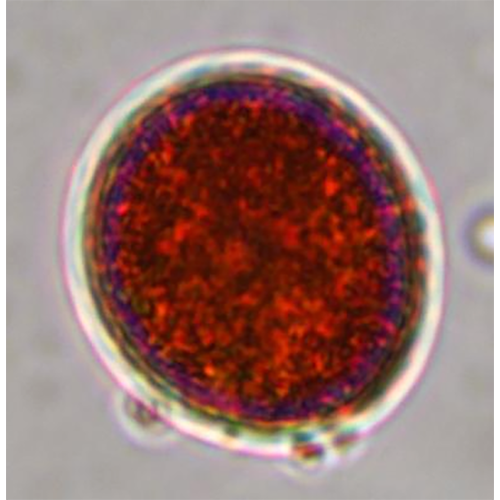
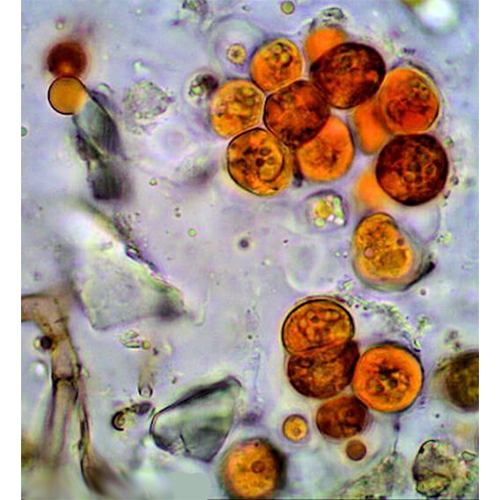
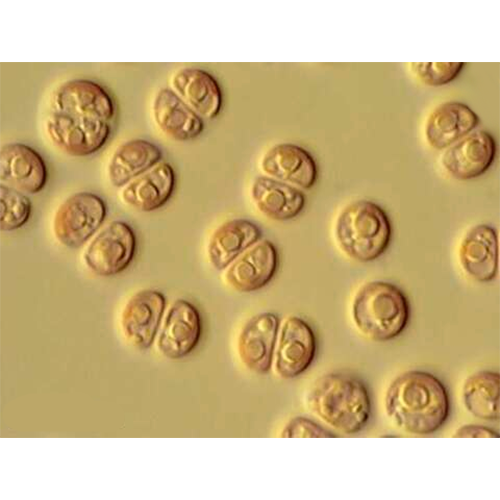

Описание:
Название происходит от греческого слова rhodon — розовый.
Красные водоросли, или багрянки (лат. Rhodóphyta) — отдел водорослей, обитатели прежде всего морских водоёмов, пресноводных представителей известно немного. Обычно это довольно крупные растения, но встречаются и микроскопические. Среди красных водорослей имеются одноклеточные (крайне редко), нитчатые и псевдопаренхимные формы; истинно паренхимные формы отсутствуют. Ископаемые остатки свидетельствуют, что это очень древняя группа растений. Красные водоросли неоднократно переходили к паразитированию на других красных водорослях: как близкородственных (адельфопаразитизм), так и более удалённых (аллопаразитизм; Harveyella mirabilis, Choreocolax polysiphoniae).
Строение
Красные водоросли — эукариоты. Хлоропласты красных водорослей двумембранные, с одиночными тилакоидами. Один или два тилакоида обычно лежат на периферии хлоропласта. На мембранах тилакоидов имеются фикобилисомы. Основным пигментом хлоропластов является хлорофилл. Кроме того, у красных водорослей имеются каротиноиды и фикобилины в фикобилисомах. Благодаря такому набору пигментов красные водоросли могут поглощать свет почти всей видимой части спектра. Как правило, хлорофилл маскируется фикобилинами (красного и синего цвета) и каротиноидами (оранжево-жёлтые), но среди пресноводных красных водорослей встречаются исключения. Так, Batrachospermum, обитающий в сфагновых болотах, сине-зелёного цвета.Запасные вещества — багрянковый крахмал (α-1,4-глюкан), запасаемый в цитоплазме, низкомолекулярный углеводород флоридозид и многоатомные спирты.
Клеточная стенка красных водорослей состоит из фибриллярного матрикса (сложен рыхло расположенными фибриллами целлюлозы или ксилана) и аморфной фракции, в состав которой могут входить агар, агароиды, каррагинаны и маннаны. У ряда красных водорослей клеточная стенка инкрустируется карбонатами кальция, магния и стронция. Иногда поверх клеточной стенки расположена белковая кутикула.
Митоз полузакрытый ацентрический. Клетки делятся за счёт впячивания клеточной мембраны. В ходе митоза образуются поры, функционально аналогичные плазмодесмам высших растений, но имеющие иное происхождение. Поры закрываются специальными поровыми пробками, состоящими из белков и полисахаридов.
У красных водорослей полностью отсутствуют жгутиковые стадии жизненного цикла.
Особенности жизненного цикла
Для красных водорослей характерен сложный цикл развития, не встречающийся у других водорослей. Репродуктивные клетки красных водорослей никогда не имеют жгутиков. Они выходят из спорангия или гаметангия в результате образования большого количества слизи и разносятся водой. Половой процесс всегда оогамный. После оплодотворения образовавшаяся зигота претерпевает сложное развитие прямо на гаметофите и даёт начало особым спорам, которые называются карпоспорами, образующиеся в карпоспорангиях, тогда как у многих других водорослей зигота развивается в спорофит, давая начало новой форме развития растений. Жизненный цикл красных водорослей изоморфный или гетероморфный дипло-гаплобионтный.Красные водоросли в морях России
В Баренцевом море красные водоросли — типичные представители прибрежной бентосной растительности. Выше других водорослей на прибойных скалах, открытых волнам, ветру и солнцу, растёт Porphyra umbilicalis. Для нижнего горизонта литорали в местах с хорошим движением воды характерен пояс литоральных багрянок, формируемый Palmaria palmata, Devaleraea ramentacea, а также Rhodomela lycopodioides, Polysiphonia urceolata и другими. На литорали, опускаясь иногда в сублитораль, растёт Chondrus crispus. В открытых местах на Мурманском побережье имеется также пояс сублиторальных багрянок, идущий глубже пояса ламинариевых водорослей (на глубине более 8 м), основными представителями которого являются: Ptilota plumosa, Odonthalia dentata, Phycodrys rubens. Глубже других идёт пояс известковых кораллиновых водорослей, это виды рода Lithothamnion sp. и некоторые другие.В Белом море на глубине 0—5 м среди фукусов и ламинарий поселяется, иногда в больших количествах, анфельция (Ahnfeltia plicata). Она нередко отрывается от субстрата и по течению сносится в бухты, где могут формироваться больших размеров не прикреплённые пласты, толщиной около 20 см. В Белом море ведётся добыча анфельции для получения агар-агара. В некоторых странах собирают Chondrus crispus, из него получают полисахарид — каррагинан.
В Чёрном море расположен заказник Малое филлофорное поле , к северо-западу от Крыма.
Палеонтология
Фосфатизированные доломитовые строматолиты в карбонатной свите Тирохан (Tirohan Dolomite) у подножья хребта Виндхья в центральной Индии, на участке реки Пайсуни в квартале Янкикунд (Jankikund) города Читракута в штате Уттар-Прадеш, возраст которых оценивается в 1,6 млрд лет, известны уникальной сохранностью окаменелых микробных образований. Палеонтологи из Шведского музея естественной истории обнаружили древнейшие окаменелые останки многоклеточных растений, возраст которых оценивается в 1,6 млрд лет. Учёные интерпретировали останки как красные водоросли. Нитевидная форма Rafatazmia chitrakootensis Bengtson состоит из одного ряда крупных клеток с ромбовидным диском внутри каждой клетки, предположительно интерпретируемым как пиреноид. Род Rafatazmia Bengtson назван в честь палеонтолога доктора Рафата Азми, Rafat Azmi, вид Rafatazmia chitrakootensis — по городу Читракута. Вторая нитевидная форма Denaricion mendax Bengtson имеет монетоподобные клетки. Название рода Denaricion Bengtson образовано от лат. denarius — денарий и греч. κίων — столб из-за структуры, похожей на стопку монет, вида — от лат. mendax — обманчивый из-за его загадочной структуры. Вид Ramathallus lobatus Bengtson имеет структуру, указывающую на родство с флоридеевыми водорослями. Название рода Ramathallus Bengtson образовано от Рама и лат. thallus — таллом, вида — от лат. lobatus — дольчатый. Исследованные окаменелые останки получены в результате полевых работ в Янкикунде в ноябре 2006 года и январе 2011 года и были дополнены окаменелыми останками, предоставленными палеонтологом Рафатом Азми. Найденные останки старше ранее найденных красных водорослей вида Bangiomorpha pubescens на 400 млн лет.Ранее найденная красная водоросль Bangiomorpha pubescens является самой древней окаменелостью-эукариотом, принадлежащей к конкретному современному таксону. Bangiomorpha pubescens, окаменелость многоклеточного организма, найденная в арктической Канаде, лишь немногим отличается от современной красной водоросли рода Бангия (Bangia), несмотря на то, что она была захоронена в отложениях возрастом 1,2 млрд лет
Использование
Некоторые виды красных водорослей употребляются в пищу. Наиболее известны среди них Palmaria palmata, грацилярия и порфира.Из красных водорослей получают гелеобразующее вещество агар-агар.
Анфельция, филлофора и другие используются в медицине.
Общая характеристика
1. Одноклеточные, колониальные и многоклеточные организмы с коккоидным, нитчатым, гетеротрихальным, псевдопаренхиматозным и паренхиматозным типами дифференциации таллома. 2. Характерно полное отсутствие жгутиковых стадий в жизненном цикле. 3. Оболочка хлоропластов образована двумя мембранами, хлоропластная эндоплазматическая сеть отсутствует. Тилакоиды одиночные, равноудаленные, не собраны в ламеллы. Хлоропластная ДНК в виде маленьких нуклеотидов, разбросанных по всей строме хлоропласта. 4. Из хлорофиллов присутствует только хлорофилл а. Дополнительные пигменты фикоэритрин, фикоцианин и аллофикоцианин собраны в фикобилисомах, имеются различные каротиноиды. 5. Основной запасной продукт — багрянковый крахмал, который откладывается в цитоплазме. 6. Митохондрии с уплощенными кристами. 7. Митоз полузакрытый, без центриолей. 8. Клеточная стенка чаше всего состоит из целлюлозы (структурная фракция) и пектиновых веществ (агар, каррагинан, агароза) аморфного матрикса. 9. Между клетками имеются поровые соединения. 10. Преобладает гапло-диплобионтный жизненный цикл с тремя многоклеточными фазами, две из которых диплоидные. 11. Обитают главным образом в морях (обычно прикрепленные формы), реже встречаются в пресных водах.Большинство красных водорослей — многоклеточные сложноустроенные организмы, и только наиболее примитивные представители имеют одноклеточное или колониальное слоевище. Форма тела багрянок весьма разнообразна: нитевидная (волосовидная или грубая), пластинчатая цельная или сложно рассеченная с дополнительными выростами по краю {папиллы — мелкие бородавчатые выросты, пролификации — более крупные, напоминающие по форме основные ветви), цилиндрическая (плотная или с полостью внутри, шнуровидная или в виде кустиков), корковидная (корки, пленки, прижатые к субстрату), коралловидная. Многообразие внешних форм сводится к нескольким типам дифференциации таллома: коккоидный, нитчатый, разнонитчатый, ложнотканевый и тканевый. Большинство красных водорослей имеет псевдопаренхиматозные талломы, возникающие за счет переплетения боковых ветвей либо одной оси, нарастающей апикально, либо многих таких осей. В первом случае говорят об одноосевом строении, во втором — о многоосевом.
Хлоропласты.
Форма хлоропластов красных водорослей может быть различной. У большинства они расположены вдоль клеточных стенок — париетально. Эти пластиды бывают дисковидными и лентовидными с многочисленными переходами между ними. Только у нескольких родов красных водорослей хлоропласты звездчатые и занимают в клетке центральное положение. Форма хлоропластов может быть разная даже у одного и того же организма. Например, у видов рода Ceramium в крупных осевых клетках междоузлий хлоропласты вытянутые, лентовидные, а в мелких клетках коры на узлах они короткие, дисковидные. Форма хлоропластов может меняться в зависимости от возраста водоросли в связи с изменениями размеров клетки, при изменении условий освещения и аэрации. Некоторые красные водоросли имеют хлоропласты с пиреноидами, функция которых не известна.
Каждый хлоропласт окружен только своей собственной двухмембранной оболочкой. Толщина каждой мембраны 13 нм. Тилакоиды, как и у глаукоцистофит, в хлоропластах не собраны в группы и лежат на одинаковом расстоянии друг от друга. Толщина каждого тилакоида около 18 — 20 нм. Один или два тилакоида расположены обычно по периферии хлоропласта параллельно его оболочке. Из хлорофиллов в пластидах красных водорослей присутствует только хлорофилл 6/, который маскируется дополнительными пигментами — красным фикоэритрином, синими фикоцианином и аллофикоцианином. Эти фикобилины локализованы на поверхности тилакоидов в фикобили- сомах полусферической и полудисковидной формы. В полусферических фико- билисомах доминируют фикоэритрины, в полудисковидных — фикоцианины. У цианобактерий фикобилисомы в основном полудисковидной формы, у красных такая форма характерна только для немногих представителей, например для Porphyridium.
Фикоэритрин у красных водорослей встречается по крайней мере в виде 5 форм (В-фикоэритрин 1 и II, R-фикоэритрин I, II и Ill), которые отличаются спектрами поглощения, хотя все имеют пик в зеленой области спектра (500 — 570 нм). Все пять типов случайно распространены среди различных групп красных водорослей. Фикоцианин встречается в форме R- и С-фикоцианинов с пиками поглощения при 615 и 620 нм, в то время как аллофикоцианин имеет пик при 650 нм. Все пики лежат в красной части спектра. Пресноводные багрянки содержат преимущественно фикоцианин, поэтому и окрашены в голубовато-зеленый цвет. Красные водоросли, которые обитают в очень сильно освещенных местах, могут быть окрашены в желтый, коричневый или черный цвет из-за присутствия в клетках большого количества различных фотозащищающих каротиноидов. Фикобилиновые пигменты водорастворимы и вымываются из клеток после отмирания водорослей.
Из дополнительных пигментов наиболее важны сх- и p-каротин, лютеин и зеаксантин. У некоторых видов обнаружены антераксантин, виолаксантин, а- и [3-криптоксантин. Хлоропластная ДНК имеет вид маленьких, 1—2 мкм в диаметре, нуклеоидов, которые разбросаны по всему хлоропласту. Возможно, что каждый нуклеоид состоит из нескольких кольцевых молекул хлоропластных ДНК. Полагают, что хлоропласты красных водорослей являются результатом первичного эндосимбиоза с цианобактерией. В то же время хлоропласты криптомонад, примнезиофит, охрофит и, возможно, большинства динофит произошли в результате вторичного эндосимбиоза путем внедрения в клетку эука- риотного хозяина клеток красных водорослей.
Запасные продукты.
Основной запасной продукт — багрянковый крахмал (а-1,4-связанный глюкан). В отличие от зеленых водорослей его зерна формируются в цитоплазме, а не в хлоропласте. Багрянковый крахмал, дающий с йодом ярко-красное окрашивание, состоит из неветвящейся амилозы, которая входит также в состав крахмала зеленых водорослей, но у него отсутствует ветвящаяся амилопектиновая фракция. Важный продукт фотосинтеза красных водорослей — низкомолекулярный углеводород флоридозид. Его содержание в талломах некоторых представителей может быть выше 10 % от сухого веса. Он выполняет осморегу- ляторную функцию. Помимо красных водорослей, флоридозид встречается у синезеленых и криптофитовых водорослей. Его концентрация в клетках увеличивается с увеличением солености среды. Некоторые красные водоросли могут запасать многоатомные спирты.
Клеточная стенка.
Клеточная стенка состоит из аморфного матрикса и фибриллярной фракции. Прочность клеточной стенке придают фибриллы из целлюлозы, которые у красных водорослей образуют неправильную сеть. У Porphyra и Bangia фибриллярная часть клеточной стенки состоит из р-1,3-связанного ксилана (полимер ксилозы), аморфная фракция состоит из маннанов, в то же время у спорофитов этих водорослей — стадия Conchocelis — фибриллярная фракция представлена целлюлозой, а аморфная состоит из галактанов. Наиболее важными веществами полисахаридного аморфного матрикса являются агар, агароиды, каррагинан. Эти вещества могут быть экстрагированы горячей водой с образованием коллоидных растворов. Синтезируются они в цистернах аппарата Гольджи, транспортируются к клеточной поверхности в везикулах, а затем встраиваются в стенку. На их долю приходится до 70% сухого веса клеточной стенки. У ряда красных водорослей поверх клеточной стенки может располагаться кутикула, состоящая главным образом из белка. Среди красных водорослей имеются представители с обызвествленными оболочками. Состав кальцинирующих соединений у них неодинаков: у одних эго — кальцит (порядок кораллиновых), у других, например у Liagora, арго- нит. Вместе с карбонатом кальция откладываются карбонаты магния и стронция. Инкрустироваться может часть таллома или весь таллом. В кальцинированных клеточных стенках некоторых красных водорослей встречается альгиновая кислота, которая входит в состав клеточных стенок бурых водорослей. Полагают, что кислые полисахариды (возможно, альгиновая кислота) могут выступать как центры кристаллизации в клеточных стенках.
Ядро.
Клетки красных водорослей обычно одноядерные, но у флоридеевых наряду с одноядерными имеются и многоядерныс. Ядра мелкие, диаметр самых мелких нс превышает 1 мкм. Размер ядер зависит от типа клетки, ее возраста и местоположения в талломе. Гаплоидный набор хромосом варьирует в пределах 2 — 68 (-72), содержание ДНК — 0,22 - 2,85 иг. Для ядер красных водорослей известна эндоредупликация (репликация ядерного генома без митоза), приводящая к полиплоидии (кратное увеличение набора хромосом в клетках). Помимо полиплоидии у красных водорослей известна и анеуплоидия (клетка содержит измененное число хромосом, не кратное гаплоидному набору).
Митоз, мейоз и цитокинез.
С помощью электронной микроскопии процессы деления ядра исследованы у ряда красных водорослей. Митоз относится к полузакрытому ацентрическому типу. На примере Polysiphonia показано, что на стадии ранней профазы на одном из полюсов будущего веретена образуется полярное кольцо в виде темноокрашенной структуры. Другое полярное кольцо мигрирует вокруг ядра к противоположному полюсу веретена. В поздней профазе полярные кольца располагаются на полюсах будущего веретена, прилегая к небольшим выпячиваниям ядерной оболочки, и окружены зоной, свободной ог рибосом. В этой зоне начинают формироваться микротрубочки. Вокруг ядра развивается перинуклеарный эндоплазматический ретикулум, ядрышко исчезает.
В метафазе хромосомы выстраиваются в мстафазную пластинку, к ним с помошыо кинстохоров прикрепляются микротрубочки веретена. Хромосомные и межнолюсные микротрубочки не сходятся в области полярных колец, поэтому полюса веретена широкие. Ядерная оболочка остается интактной, хотя и перфорирована крупными порами; на полюсах формируются отверстия. Между ядром и перинуклеарным эндоплазматическим ретикулумом располагаются перинуклеарные микротрубочки.
Межполюсныс микротрубочки существуют во время поздней анафазы и ранней телофазы; тслофазное веретено удлиняется. В поздней телофазе между образовавшимися ядрами закладывается вакуоль.
Цитокинез идет за счет виячивания цитоплазматической мембраны. На месте вакуоли формируется септальная пора, которая закрывается белковой поровой пробочкой.
Митоз у одноклеточных красных водорослей имеет ряд особенностей: отсутствие перинуклеарной оболочки, образование очень широких полярных отверстий, появление в поздней анафазе характерной межзональной зоны, слабая дифференциация кинетохоров и прикрепление к ним только одной микротрубочки.
Мсйоз у красных водорослей доказан цитологически по наличию синаптонемного комплекса в молодых тетраспорангиях. Синаптонсмный комплекс имеет такую же структуру, как и у других эукариот. Он состоит из длинной центральной части, к которой с каждой стороны прикрепляются гомологичные хромосомы. Центральная часть белковая и состоит из центрального элемента, с каждой стороны которого располагаются в два ряда поперечные элементы. С другой стороны поперечных элементов находятся боковые элементы, к которым прикрепляются петли хроматина хромосом. Концы боковых элементов прикреплены к внутренней мембране ядерной оболочки.
Поровые соединения.
Наличие поровых соединений — отличительная особенность красных водорослей, имеющая значение для выделения порядков. Встречается два типа поровых соединений — первичное и вторичное. Первичное поровое соединение формируется между двумя дочерними клетками при делении материнской, вторичное — при слиянии двух клеток. В образовании пробочек участвует эндоплазматическая сеть. Ее цистерны выстраиваются в септальной поре, в них и снаружи от них откладывается материал пробки. Внутри поровые пробочки заполнены белковым содержимым, шапочки частично состоят из полисахаридов.
По крайней мере существует 7 различных типов поровых пробок.
- Тип 1. Пробочки окружены с каждой стороны двухслойными шапочками. Наружный слой шапочки отделен от внутреннего слоя шапочковой мембраной, которая соединена с плазмалеммой. Встречается у немалиевых, пальмарисвых, акрохстисвых.
- Тип 2. В отличие от типа 1 наружный слой шапочки утолщен. Встречается у батрахоспермовых, родогоргоновых.
- Тип 3. В отличие от типа 2 отсутствуют шапочковые мембраны. Встречается у порядка кораллиновые.
- Тип 4. Поровые пробочки окружены однослойными шапочками, имеется шапочковая мембрана. Встречается у порядка гелидисвые и гильденбрандиевые.
- Тип 5. В отличие от типа 4 отсутствуют шапочковые мембраны. Встречается у бангиевых.
- Тип 6. Шапочки отсутствуют. Пробочки окружены с каждой стороны шапочковой мембраной. Встречается у гигартиновых, родимениевых, церамисвых, боннемезонисвых.
- Тип 7. В отличие от типа 6 отсутствуют шапочковые мембраны. Встречается у родохетовых, анфельциевых.
Размножение.
Размножение у красных водорослей — вегетативное, бесполое и половое. Половое воспроизведение оогамное, имеет ряд особенностей, не встречающихся у других водорослей. Мужские половые клетки — спермации, лишены жгутиков и пассивно с током воды переносятся к женским половым органам — карпогонам. После оплодотворения формируются нити гонимобластов, клетки которых превращаются в карпоспорангии. Совокупность гонимобластов называется гонимокарпом (в литературе часто используют термин карпоспорофит). У ряда флоридей гонимокарп окружает оболочка. Тогда эта структура называется цистокарпий. В карпоспорангиях образуются карпоспоры, которые прорастают в диплоидный тстраспорофит. На тетраспорофите в тетраспорангиях происходит редукционное деление. Гаплоидные тетраспоры прорастают в гаплоидный гаметофит. Этот жизненный цикл с чередованием поколений (одного гаплоидного — гаметофит и двух диплоидных ~ карпоспорофит и тетраспорофит) встречается у большинства красных водорослей, но существует ряд отклонений от него. Происхождение такого уникального жизненного цикла — эволюционная компенсация за потерю жгутиков.
Тетраспоры у красных водорослей могут располагаться в тетраспорангиях по-разному. У наиболее эволюционно продвинутых представителей они находятся в углах тетраэдра, у других могут располагаться друг над другом — линейно (зонально). Известно крестообразное расположение и крестообразное с поворотом двух нижних тетраспор относительно верхних.
Карпогон красных водорослей имеет вид колбы и состоит из расширенной нижней части (брюшка) и вытянутой верхней — трихогины, для улавливания спсрмациев. В карпогонс обычно два ядра: одно, лежащее в трихогине, быстро дегенерирует после созревания карпогона, второе расположено в базальной части карпогона и функционирует как ядро женской гаметы. У большинства красных водорослей карпогон формируется на конце короткой, часто ветвящейся, трех- или чстырехклсточной ветви, называемой карпогонной. Клетка, дающая начало карпогонной ветви, называется поддерживающей. Карпогон и карпогонная ветвь обычно бесцветные, за исключением нсмалиевых.
Спермации формируются по одному в сперматангиях, которые в свою очередь образуются на материнских клетках. В молодых сперматангиях ядро расположено в апикальной части, в то время как одна или больше вакуолей — в базальной. При созревании сперматангиев вакуоли сливаются в одну крупную, занимающую половину объема клетки. В вакуоли содержится фибриллярный материал (возможно, мукополисахариды), выделение которого обеспечивает выход спюрмация через желированный участок клеточной стенки около верхушки. Зрелый спермапий одноядерный и лишен жесткой клеточной стенки, окружен слизыо, может содержать хлоропласты.
Спермаций пассивно переносится токами воды, контактирует с трихогиной, которая расположена над поверхностью женского гаметофита. Стенки спсрмацисв и трихогины в месте контакта растворяются, мужское ядро проходит через центральный канал в трихогине и сливается с гаплоидным ядром карпогона. Далее развитие гонимокарпа у различных красных водорослей (карпоспорофита) имеет свои особенности. Во-первых, нити гонимобласта могут развиваться непосредственно из брюшка карпогона. Во-вторых, у большинства красных водорослей гонимобласты развиваются из особых вспомогательных вегетативных клеток — ауксиллярных, которые могут быть или удалены от брюшка карпогона или располагаются в непосредственной близости от него. Если ауксиллярные клетки удалены от карпогона, из его брюшка развиваются диплоидные ообластемные (соединительные) нити. Они дорастают до ауксиллярной клетки, происходит плазмогамия без кариогамии, и это слияние стимулирует образование нитей гонимобластов. Если ауксиллярная клетка формируется в непосредственной близости к карпого- ну и только после его оплодотворения, то такая совокупность этих клеток называется прокарпии. Ауксиллярная клетка здесь просто сливается с брюшком оплодотворенного карпогона, после чего развиваются гонимобласты с карпоспорами.
Экология и значение.
Подавляющее большинство известных красных водорослей — обитатели морей, причем встречаются они во всех морях Мирового океана и лишь около 200 видов в пресных водоемах: реках, прудах, озерах, лужах. Благодаря дополнительным красным пигментам красные водоросли могут расти на значительных глубинах (100 — 200 м) при наличии подходящего грунта и хорошей прозрачности воды. Чаще всего их заросли кончаются на глубине 20 — 40 м. Обильно они представлены на литорали. Некоторые багрянки приспособились к жизни в зоне заплссков и брызг. Среди красных водорослей есть небольшое число наземных обитателей.
В морях красные водоросли встречаются повсеместно. Они поселяются на скалах, валунах, каменистых россыпях, на искусственных сооружениях, других растениях и на панцирях морских животных. Многие багрянки растут как эпифиты на других водорослях, в том числе красных, т.е. они используют другие водоросли только как субстрат для прикрепления. Среди эпифитов встречаются облигатные формы, например Polysiphonia lanosa, которая обитает на бурой водоросли Ascophyllum. После прорастания споры Polysiphonia на поверхности хозяина развивается ризоид, который закрепляется в тканях хозяина, переваривая рядом лежащие ткани с помощью ряда ферментов, освобождающихся из везикул на конце ризоида. Polysiphonia — облигатный эпифит. В отличие от паразитов у нес нс существует переноса метаболитов от хозяина, она сама обеспечивает себя питательными веществами за счет фотосинтеза.
В то же время немало багрянок ведут паразитический и полупаразитичский образ жизни, они не только погружены в ткани хозяина, но и питаются за его счет. Среди красных водорослей встречаются аделъфопаразиты (паразиты, которые тесно в таксономическом смысле связаны со своим хозяином) и аллопаразиты (паразиты, которые не близки своему хозяину). Адельфопаразиты встречаются среди представителей порядков грациляриевых, гигартиновых, плокамисвых, родимсниевых. С помощью методов геносистематики показано, что каждый такой паразит эволюционировал или непосредственно от хозяина, на котором в настоящее время он паразитирует, или от других таксонов, тесно связанных с современным хозяином. Адсльфопаразиты легче устанавливают вторичное поровое соединение с клетками хозяина, что позволяет переносить питательные вещества.
Красные водоросли играют значительную роль в морских экосистемах. Они — продуценты первичной продукции и местообитание для других морских организмов. Красные водоросли из порядка кораллиновые принимают участие в образовании коралловых рифов. Рифы строятся в результате роста кораллов и кораллиновых красных водорослей (главным образом виды Porolithon).
Красные водоросли широко используются человеком в хозяйстве и в быту. В качестве пищевых продуктов в мире используют некоторые виды Bangia, Porphyra, Dermonema, Asparagopsis, Grateloupia, Gigartina.
Красные водоросли — продуценты сульфатированных полигалактанов (агара, агарозы, каррагинана). Агар получают из водорослей — агарофитов (Gelidiит, Gracilaria, Pterocladium), Основной производитель агара в настоящее время — Япония, в которой его производят с XVII в. Япония и Корея ежегодно получают 3 500 т агара, что составляет половину всей мировой продукции. В России в 90-х гг. XX в. производилось 1 200 т агароидных веществ, из них чистого агара — 300 т; производство высококачественного агара составляло всего 100 т. В российских морских водах нет промышленных зарослей хороших агарофитов, поэтому на Дальнем Востоке и Белом море налажсно производство агара из Ahnfeltia, в южном Приморье ддя этих целей используют также Gracilaria. Агар имеет большое значение в микробиологических исследованиях. Питательные среды, содержащие 1 — 2% агара, используют для роста бактерий и грибов; этот гель устойчив сам по себе к деградации этими организмами. Кроме того, агар нс токсичен, поэтому его активно используют в пищевой промышленности (для получения мармелада, джема, майонезов, консервирования мяса и рыбы).
Показана антикоагулянтная активность агара, однако механизм действия значительно отличается от того, который известен для гепарина. Применяют агар для лечения ожогов, в стоматологии, при изготовлении капсул для лекарственных препаратов.
Широко используют агар в различных отраслях промышленности не только как желирующее средство, но и как эмульсионный и связующий агент. Агароза используется в молекулярной биологии в качестве молекулярного сита для разделения высокомолекулярных соединений.
Каррагинаны получают из Chondrus crispus, Mastocarpus stellatus; на Филлипинах для этих целей специально культивируют Eucheuma и Kappaphycus. В год получают около 13 тыс. т каррагинанов, из них более половины в Европе и около половины на Филиппинах и в Индонезии. Каррагинан не токсичен, поэтому используется в пищевой промышленности при изготовлении желированных продуктов, мороженого, майонеза и др., в парфюмерии — для производства кремов, зубных паст, в фармацевтической и текстильной промышленности — для стабилизации эмульсий и суспензий. Он обладает хорошо выраженным противовирусным, антикоагуляционным и противоязвенным действием.
Филогения.
Вопрос о происхождении и филогенетических связях красных водорослей с другими организмами до сих пор трактуется разными авторами по-разному. Красные водоросли — древняя группа организмов. Сообщается об ископаемых формах, датированных возрастом 1,9 млрд лет, которые интерпретируются как Porphyridium-подобные родофиты. Нахождение многоклеточных ископаемых таксонов Bangiomorpha (1,2 млрд лет) представляет собой наиболее раннюю находку среди многоклеточных эукариот с половым процессом.
Среди красных водорослей окаменелые остатки известны для кораллино- вых водорослей. Представители семейства Solenoporaccac (полностью вымерли) впервые появились в кембрии и были обильны в ордовике. Нечленистые кораллиновые водоросли в виде свободноживущих структур известны в ископаемом и современном состоянии как родолиты. Они широко распространены на дне океанов на глубине от 50 до 200 м, обладают медленным ростом и за 800 и более лет их диаметр может достигать 30 см. Возраст некоторых родолитов, определенный радиоуглеродным методом, составляет более 13 тыс. лет.
Окаменелости некальцинированных красных водорослей представлены плохо, хотя известны. Имеются достоверные находки таких водорослей из верхнего силура.
В последние годы красные водоросли все чаше рассматривают как сестринскую группу зеленых растений, обосновывая это доказательствами как на макроуровне (особенности строения), так и на молекулярном уровне. Что касается филогенетических связей в рамках красных водорослей, то на ультраструктурном и молекулярном уровнях показано, что бангисвые (в традиционном понимании этого класса) — парафилетичная группа, в то время как флориде- евые — монофилстичная. Характерным для бангисвых долгое время считалось отсутствие поровых соединений, апикального роста и полового размножения, в то время как флоридиевые имели поровые соединения, апикальный рост и трехфазный жизненный цикл. Однако у ряда бангиевых (родохетовые, бангисвые) также были обнаружены поровое соединение, апикальный рост и половое размножение, в то же время не у всех флоридиевых имеется апикальный рост (например, отсутствует у кораллиновых), не у всех существует трехфазный жизненный цикл (например, у батрахоспермовых). Сравнение последовательностей нуклеотидов ряда генов убедительно показало, что красные водоросли можно разделить на две группы: одна включает только цианидие- вых, а другая — всех остальных красных водорослей. Ясно также показано, что только порядок бангиевых из традиционного класса бангиофициевых формирует сестринскую группу для флоридей. Остальные бывшие бангиофицисвые формируют самостоятельные группы.
Книги:
 Ботаника, Том 1, Водоросли и грибы, Белякова Г.А., Дьяков Ю.Т., Тарасов К.Л., 2006
Ботаника, Том 1, Водоросли и грибы, Белякова Г.А., Дьяков Ю.Т., Тарасов К.Л., 2006
 Ботаника, Том 2, Водоросли и грибы, Белякова Г.А., Дьяков Ю.Т., Тарасов К.Л., 2006
Ботаника, Том 2, Водоросли и грибы, Белякова Г.А., Дьяков Ю.Т., Тарасов К.Л., 2006
 Фёдоров А.А. Жизнь растений в шести томах. Том 3. Водоросли. Лишайники
Фёдоров А.А. Жизнь растений в шести томах. Том 3. Водоросли. Лишайники
 Забинкова Н. Н., Кирпичников М. Э. Латинско-русский словарь для ботаников. — М., Л.: Изд-во АН СССР, 1957. — 335 с.
Забинкова Н. Н., Кирпичников М. Э. Латинско-русский словарь для ботаников. — М., Л.: Изд-во АН СССР, 1957. — 335 с.