Описание:
Класс брюхо ресничных содержит несколько сотен видов микроскопических ресничных червеобразных существ, обычных в пресных водах и в море. Природа и филогенетическое положение этих организмов были выяснены не сразу. Их первые изображения (под наименованием «инфузории») принадлежат натуралисту XVIII в. О.Ф.Мюллеру. Уже в XIX в. известный микроскопист Эренберг описал и ввел в науку первые роды пресноводных гастротрих Chaetonotus и Ichthydium, но обозначил их коловратками. В середине прошлого века Ш у л ь ц е открыл первую морскую гастротриху Turbanella и выдвинул идею близости гастротрих к турбелляриям.
Собственно название Gastrotricha (т. е. буквально «брюхоресничные») принадлежит нашему соотечественнику И.И.Мечникову. Руководствуясь идеей о родстве гастротрих и коловраток он объединил эти две группы в один класс. Людвиг был первым, кто нашел в организации гастротрих черты, объединяющие их с нематодами. Чрезвычайно много занимался гастротрихами Зелинка; в 1899 г. он опубликовал монографию, суммировавшую все известные к тому времени данные по анатомии и систематике гастротрих. Зелинка вновь объединил гастротрих и коловраток, введя для них группу Trochelminthes на том основании, что, по его мнению, те и другие соответствуют по строению трохофоре — плавающей ресничной личинке многощетинковых червей и некоторых моллюсков. В первой половине XX в. наибольший вклад в познание гастротрих сделал крупнейший современный немецкий зоолог А. Р е м а н е, известный, в частности, своими замечательными работами по так называемым интерстициальным животным — микроскопическим обитателям интерстициев — капиллярных пространств между зернами песка на морских пляжах. Монография Ремане до сих пор служит справочным источником по этой группе животных. Наконец, совершенно новый период изучения гастротрих открыли тонкие морфологические работы, сделанные в 1960—1970-х годах с применением электронной микроскопии западноевропейскими и американскими учеными Г. Тойхерт, Е. Руппертом, Р. М. и Г. Е. Ригерами и др. Помимо чисто анатомических, в наше время интенсивно развиваются исследования по экологии и биологии гастротрих, особенно морских литоральных форм; ведь гастротрихи — типичные животные интерстициальной фауны.
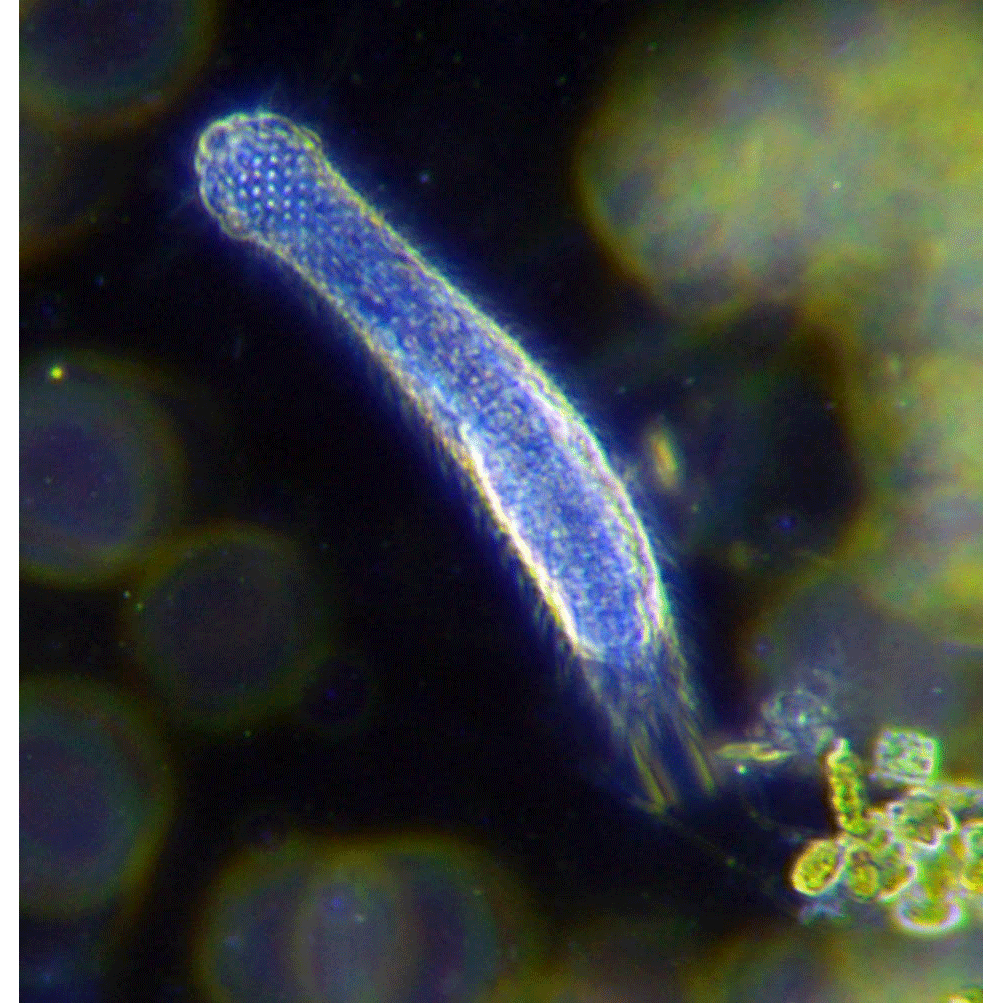
Строение. В классе гастротрих два отряда: хетонотиды (Chaetonotida) объединяют два пресноводных, два морских и два смешанных семейства; макродазиды (Macrodasyida) содержат морские формы (несколько семейств), проникающие изредка и в опресненные воды. Тело гастротрих из отряда Chaetonotida короткое, по очертаниям чаще похоже на бутылку — с шейным перехватом и расширенной задней частью. Гастротрихи Macrodasyida обычно удлиненные равномерной ширины, без перехватов и вздутий. На поперечном срезе тело гастротрих сплющенное, точнее треугольное, с широкой ползательной брюшной стороной. На переднем конце тела рот, на заднем анальное отверстие, слегка смещенное на брюшную сторону. Основной орган передвижения гастротрих — мерцательный эпидермис. Однако сплошного ресничного покрова, как, например, у турбеллярий, у гастротрих нет, локомоторные (движущие) реснички сосредоточены на брюшной стороне тела (отчего этих червей называют еще брюхоресничными). Реснички в примитивном случае представлены сплошным брюшным полем, тянущимся от головы до хвоста, но у многих родов сохраняются только в головном отделе или распадаются на отдельные продольные или поперечные мерцательные полосы и пучки. Интересно, что в одной группе морских гастротрих — хетонотид (Xenotrichulidae) реснички организованы в плотные пучки — ц и р р ы, каж у инфузорий Hypotricha. Сравнительна недавно обнаружилось, что многие гастротрихе имеют так называемый моноцил парный, или жгутиковый, эпидермис. Моноцилпарным называют такой эпидермис, где на одну покровную клетку приходится одна-единственнаг ресничка, называемая в таком случае жгутом. На ультраструктурном уровне жгуты моно-цилиарного эпидермиса не отличаются по строению от ресничек более обычного для многоклеточных животных полицилиарного эпидермиса (где на одну клетку приходится много ресничек Поэтому, строго говоря, разница между двум типами мерцательных эпидермисов невелика. Однако жгутиковый эпидермис эволюционно более древний. У современных Metazoa, кроме гастротрих, им обладает только трихоплакс (тип Placozoa) — живая модель предка всех многоклеточных, да еще гнатостомулиды — оригинальные микроскопические турбелляриеобразные черви.
Все тело гастротрихи покрыто тонкой эластичной кутикулой, причем тончайшая кутикула покрывает и реснички. У многих родов, в первую очередь у хетонотид, кутикула спинной стороны тела образует разнообразные «украшения» — ши-пики, часто сложной формы и чешуйки. Чешуйки могут образовывать сплошной покров спинной стороны, причем чешуйки не прижаты непосредственно к эпидермису, а сидят на коротких стебельках, растущих от расширенных чешуевидных оснований. Таким образом, гастротриха имеет два плотных чешуйчатых покрова с пространством между ними.
Чрезвычайно характерны для гастротрих специальные органы — прикрепительные трубочки. У хетонотид они помещаются в хвостовой вилке, у макродазид — сериально по боковым краям тела. Посредством этих органов гастротрихи могут прикрепляться к субстрату, а морские интерстициальные макродазиды даже «шагать» по песчинкам, напоминая способом движения пиявок или гусениц пядениц. Часто у макродазид из трубочки торчит длинный неподвижный жгутик — чувствительный тактильный орган. В таком случае внутри каждой прикрепительной трубочки лежат три клетки: железистая,, выделяющая клейкий секрет, которым весь орган крепится к субстрату; чувствительная со жгутиком; поддерживающая, внутри которой имеются два опорных стержня, а также капли секрета, но другого свойства, чем в железистой клетке. Тонкое строение прикрепительной трубочки изучено у макродазиды Turbanella сог-nuta. Интересно, что подобное соединение чувствительной жгутиковой и железистой клеток характерно для нематод, которые многими зоологами принимаются за филогенетических потомков гастротрих или гастротрихоподобных организмов. Железистая клетка прикрепительной трубочки продуцирует клейкое вещество, а поддерживающая — его растворитель, нужный для открепления трубочки от субстрата. Подобным образом действуют и органы прикрепления — хвостовые железы — примитивных водных свободноживущих нематод. Помимо прикрепительных трубочек, в эпидермисе есть и другие одноклеточные железы разного назначения.
Ротовое отверстие ведет в сильную мускулистую глотку — фаринкс, передняя часть которой нередко расширена в небольшую ротовую полость. Фаринкс состоит из эпителиальных клеток, накрепко соединенных между собой взаимопроникающими отростками. В клетках фаринкса густыми пучками расположены сократимые мышечные волокна — миофибриллы; расположены они настолько густо, что клеточные ядра деформированы. Во внутреннем просвете фаринкса поверхность эпителиальных клеток покрыта продолжающейся внутрь тонкой кутикулой, снаружи фаринкс окружен базальной мембраной. У некоторых родов задняя часть глотки сильно утолщена,, вероятно, для усиления работы всего органа. У всех гастротрих внутренний просвет глотки на поперечном срезе треугольный, только в отряде Chaetonotida треугольный просвет сориентирован, как у нематод, основанием к спинной стороне, вершиной к брюшной, а у Macrodasyida, наоборот,— вершиной к спинной стороне, основанием к брюшной. Только у Macrodasyida глотка имеет образование, уникальное для низших беспозвоночных,— пару боковых сквозных каналов с порами, через которые полость глотки сообщается с внешней средой. Через специальный клапан глотка сообщается с кишкой короткой прямой трубкой, образованной одним слоем эпителиальных клеток. Средняя кишка переходит в заднюю кишку, кончающуюся анальным отверстием. Задняя кишка с кутикулярной выстилкой имеется не всегда — наружу может открываться непосредственно средняя кишка. Покровы гастротрих — клеточный или синцитиальный (т. е. без клеточных границ) эпидермис, образующий утолщения по бокам тела с брюшной стороны.
Нервная система представлена головным «мозгом», окологлоточным нервным кольцом и двумя продольными боковыми нервами, проходящими в боковых утолщениях эпидермиса. «Мозг» гастротрих — масса нейронов, окружающих глотку спереди и сзади от окологлоточного нервного кольца. Специальной клеточной оболочки у «мозга» нет, он окружен чувствительными и эпи-дермальными клетками. В разных его участках находятся различные типы нейронов: чувствительные, двигательные, ассоциативные и т. д. Окологлоточное кольцо и продольные нервы образованы безъядерными отростками нервных клеток, тел нейронов в них нет или очень мало. Эфферентные волокна связаны с железами и мышечными клетками — последние дают к нервам короткие синаптические отростки. Органы чувств гастротрих разнообразны. Во-первых, это специализированные покровные клетки; на одном, поверхностном конце такой клетки неподвижный чувствительный жгутик, на другом отросток — нейрит, идущий к боковому нерву. Такие тактильные рецепторы есть не только на поверхности тела, но и в просвете глотки. У многих гастротрих описаны глазки, встроенные непосредственно в поверхность «мозга». Наконец, характерны для гастротрих боковые ресничные ямки. Они располагаются позади глазков и, вероятно, являются органами химического чувства — хеморецепторами. Ресничная ямка представляет собой углубление, заполненное слизью, в котором помещаются неподвижные сложноразветвленные жгуты чувствительных клеток, соединенных нейритами с «мозгом».
У гастротрих есть и кольцевая, и продольная мускулатура, однако она не образует такого сильного и равномерного кожно-мышечного мешка, как, например, у турбеллярий. У сравнительно примитивных морских макродазид, например у Turbanella cornuta, кольцевые мышечные клетки образуют два сплошных слоя: обкладку эпидермиса изнутри и обкладку кишки снаружи. Продольные мышцы лежат двумя мощными лентами вдоль боковых сторон тела внутри от кольцевых мышц. У многих макродазид кольцевая мускулатура на отдельных участках редуцируется, а у хетонотид, видимо, совершенно отсутствует. По тонкой структуре мышцы гастротрих можно классифицировать как поперечнополосатые (глоточные и туловищные) и косо-исчерченные (туловищные). Известно, что способность мышцы к быстрому сокращению связана со сложностью ее строения. Видимо, поэтому глотка, интенсивно работающий орган, имеет поперечнополосатые волокна. Интересно, что у разных видов Macrodasyida тип туловищной мускулатуры можно связать с образом жизни животных: формы, быстро двигающиеся по поверхности грунта (Dendrodasys gracilis, Paraturbanella dohrni. Dactylopodalia baltica), имеют быстро сокращающиеся поперечно-полосатые мышцы тела, а, например, Turbanella cornuta, живущая в более спокойных условиях интерстициали в глубине грунта, снабжена косо-исчерченной туловищной мускулатурой.
У макродазид семейства турбанеллид (Turba-nellidae) и, возможно, у некоторых других есть специфическое образование — так называемый Y-орган. В типичном случае он представляет собой два тяжа очень крупных, сильно вакуолизированных клеток по обе стороны от глотки и кишки. Оба тяжа соединяются перемычками спереди, перед «мозгом», и сзади, позади ануса. Видимо, Y-орган — гидроскелет. Дело в том, что у гастротрих нет полости тела — свободного пространства между внутренними органами, заполненного жидкостью под давлением. Опорную функцию полости тела берет на себя Y-орган: у гастротрих эта упругая струна — антагонист продольной мускулатуры. Турбанеллиды, снабженные Y-органом, могут изгибать тело так же, как это делают нематоды. Гастротрихи, не имеющие Y-органа, например червеобразные Масгоiasvs, не сгибаются, но зато сильно сокращаются и вытягиваются при движении, очевидно, благодаря кольцевым мышцам. Сравнительно недавно открыта удивительная макродазида Chordodasys, жмеющая даже «хорду» — стопку вакуолизирован-ных мышечных клеток, примыкающих к кишечнику сзади в районе анального отверстия. Гистологически этот орган, функция которого не вполне ясна, действительно похож на хорду ланцетника. У гастротрих встречаются и другие хордоподобные образования, имеющие, очевидно, опорную функцию, например вакуолизированный эпидермис Macrodasys или вакуолизированные отростки мышечных клеток Dactylopodalia baltica. Опорными образованиями хетонотид, видимо, служат кутикулярные шипы и чешуйки, образующие в соединении друг с другом жесткий каркас.
Органы осморегуляции и выделения гастротрих — протонефридии — специфические ресничные органы, широко распространенные у низших червей. Основное назначение протонефридия — выводить из тела избыточную воду, которая непрерывно поступает внутрь через покровы червя из внешней среды под действием сил осмоса. Недавно строение протонефридия макродазиды изучено на примере Turbanella cornuta. Начальный элемент протонефридия — жгутиковые клетки циртоциты. Циртоцит имеет двойной цилиндр, в котором непрерывно бьется жгутик. Наружная стенка цилиндра — плазматическая манжета с порами и щелями, внутренняя образована тесно переплетенными фибриллами и поддерживается плазматическими опорами. Внутренняя жидкость проходит через двойной цилиндр, как через двойной фильтр, и биением жгутика направляется внутрь большой коллекторной клетки. В нее открываются несколько циртоцитов. В цитоплазме коллекторной клетки сквозных каналов нет, выделяемая жидкость собирается в вакуоли, которые проходят дополнительную обработку и в конце концов подходят к тому участку, где к коллекторной клетке пристроен канал, образованный единственной выводной клеткой. Биением жгутика, принадлежащего коллекторной клетке, жидкость выводится через этот канал наружу. Протонефридии Chaetonotida устроены несколько иначе и попроще. Казалось бы, именно пресноводные хетонотиды должны иметь мощную протонефридиальную систему — ведь они живут в среде, где концентрация солей резко понижена по сравнению с содержанием солей в теле, тогда как для морских макродазид разница в концентрации растворенных веществ среды и тела невелика. Однако все дело в том, что макродазиды живут не просто в море, а на литорали — прибрежной регулярно осушаемой полосе, где в отлив сильное испарение может вызвать повышение солености, а дождь — сильное распре-снение. В такой среде с резко меняющейся соленостью животные не могут обойтись без органов, обеспечивающих солевой гомеостаз их внутренней среды. Как и другие обитатели литорали, макродазиды приспособлены к колебаниям солености, т. е. являются эвригалинными формами.
Размножение. Морские гастротрихи — гермафродиты. Обычно гонады вытянуты вдоль тела справа и слева от кишки. Иногда спереди размещаются два семенника и сзади два яичника, иногда яичник один непарный. В ряде случаев гастротрихи имеют только пару гонад двойственной природы: в передней части гонады делятся и отделяются клетки, которые потом превращаются в сперматозоиды, а в задней части гонады образуются овоциты, далее превращающиеся в яйца. Эпидермальная цельная оболочка гонад не выражена — половые клетки окружены кольцевыми мышцами, клетками Y-органа, единичными мезо-телиальными клетками. Протоки половой системы выражены очень слабо. Например, настоящего яйцевода может не быть, в таком случае яйцо выходит наружу непосредственно через разрыв в стенке тела. Чаще всего у гастротрих нет специального копулятивного органа (пениса), что отличает их от турбеллярий, однако оплодотворение тем не менее внутреннее. Оплодотворение совершается с помощью сложного так называемого каудального органа, одна часть которого действует как бурса — приемник спермы, другая как пенис. Для некоторых гастро-трих-макродазид описано довольно сложное брачное поведение, включающее «роение» перед спариванием и последовательные этапы собственно спаривания.
Долгое время считали, что пресноводные хетонотиды представлены только самками, размножающимися партеногенетически, подобно коловраткам бделлоидеям. Партеногенетические самки откладывают яйца двух родов — с у б и т а н-н ы е, начинающие развиваться немедленно после откладки,; и латентные, с плотной оболочкой, хорошо переносящие высыхание и замораживание. Обычно латентные яйца откладываются в конце лета и развиваются после зимнего периода покоя. Все гастротрихи откладывают небольшое число крупных яиц; особенно велики яйца пресноводных Chaetonotida: по относительному размеру яиц сравнительно с длиной тела эта группа едва ли не первая в животном царстве. Недавно сделано открытие: некоторые пресноводные хетонотиды, например представители рода Chaetono-tus, размножаются только в том случае, если в сосуде присутствует не менее двух особей, что само по себе предполагает существование полового процесса. Почти одновременно выяснилось, что в природе имеются особи-гермафродиты со спермиями; их немного (около 8% популяции), к тому же их относительное количество может резко меняться. Видимо, пресноводные гастротрихи все же обычно размножаются партеногенетически, а гермафродитизм — временное, преходящее состояние.
Распространение. Пресноводных гастротрих можно обнаружить в разных водоемах: в прудах, на дне или на листьях погруженных растений, в выжимках болотного мха-сфагнума, где наряду с гастротрихами во множестве обитают раковинные амебы, инфузории, коловратки Bdelloidea, нематоды и тихоходки. Хетонотиды семейств Neogosseidae и Dasyditidae ведут полупелагический образ жизни. Они снабжены длинными шипиками, видимо, облегчающими парение в толще воды, а их прикрепительные трубочки редуцированы за ненадобностью. В море живут как Chaetonotida, так и Macrodasyida. Если первые чаще живут на поверхности субстрата (например, грунта), то вторые — типичные обитатели капиллярных пространств между песчинками (интерстициев) на морских пляжах. Макродазиды имеют все особенности интерстициальных животных: длинное червеобразное тело облегчает движение по тесным щелям между зернами песка, сильно развитыми органами прикрепления (прикрепительные клейкие трубочки) гастротрихи надежно приклеиваются к песчинкам. Оплодотворение внутреннее, развитие прямое, без пелагической личинки. Некоторые макродазиды имеют причудливые выросты с чувствительными жгутиками на них, некоторые, например живородящая Urodasys, снабжены длинными хвостами. Несмотря на отсутствие расселительной стадии, многие виды морских интерстициальных гастротрих распространены чрезвычайно широко, заселяя в разных морях и океанах один и тот же биотоп — интер-стициаль песчаных пляжей. Нередко на одном пляже обитает много форм гастротрих. В таком случае отдельные виды распределены по пляжу не случайно: каждый вид населяет свою зону литорали по горизонтали и свой диапазон глубин в толще песка; некоторые проникают на глубину нескольких десятков сантиметров под поверхность песка. Так разные виды гастротрих достигают пространственного разобщения и уходят от конкурентной борьбы, что особенно важно для близких видов.
Питание. Питание гастротрих изучено еще плохо. В основном они потребляют бактерий, простейших, одноклеточные водоросли, в том числе диатомовые. Иногда гастротрихи узко приспособлены к питанию каким-нибудь одним объектом. Например для интерстициальной макродазиды Turbanella cornuta показано, что она живет исключительно за счет одного-единственного вида одноклеточных водорослей — динофлагеллят.
Брюхоресничные черви, или гастротрихи (лат. Gastrotricha) — тип первичноротых животных (Protostomia). Мелкие (0,1—1,5 мм) морские и пресноводные, преимущественно донные животные, большинство видов обитает в интерстициали. Около 350 морских и около 250 пресноводных видов. / (Википедия)
Книги:
 Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987
Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987
 Зоология беспозвоночных в двух томах. Том 2: от артропод до иглокожих и хордовых / Вестхайде В., Ригер Р — 2008 с
Зоология беспозвоночных в двух томах. Том 2: от артропод до иглокожих и хордовых / Вестхайде В., Ригер Р — 2008 с
 Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989
Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989