




Описание:
Трематоды — паразитический класс плоских червей. Сохраняя все основные признаки типа, они характеризуются специфическими особенностями, которые обусловлены их эндопаразитическим образом жизни. Прежде всего это наличие специальных органов прикрепления — присосок и замена ресничного эпителия лишенным ресничек т е г у-ментом.
Пищеварительная система трематод, как правило, развита хорошо, но имеется тенденция к ее рудиментации и исчезновению. Одновременно с этим приобретает особо важное значение способность трематод к восприятию пищевых веществ извне, через покровы. Как и другие паразитические организмы, трематоды разных поколений производят огромное количество яиц и личинок. Трематодам присуща также тенденция к живорождению, полностью осуществленная на некоторых этапах их жизненного цикла.
Жизненный цикл трематод. Сам по себе жизненный цикл трематод, связанный с чередованием поколений, совершенно не свойствен свободножи-вущим плоским червям и возник в ходе эволюции, как следствие перехода этой группы к паразитизму.
Жизненный цикл трематод представляет собой одно из самых удивительных явлений живой природы. Он очень сложен, проходит с участием нескольких хозяев, сопровождается чередованием поколений и сменой паразитических и свободноживущих фаз развития.
Половозрелая гермафродитная особь — м а р и-т а паразитирует во внутренних органах позвоночных животных. Откладываемые ею яйца выводятся из организма хозяина во внешнюю среду. Это сложные яйца, под плотной скорлупкой которых находится не только яйцеклетка, но и множество желточных клеток, поставляющих необходимые развивающемуся зародышу питательные вещества. В типичном случае яйцо для завершения своего развития должно попасть в воду, и там из него выходит подвижная личинка - ми-рацидий. За время короткой жизни эта личинка должна проникнуть в первого промежуточного хозяина — моллюска определенного вида, в котором претерпевает превращение (метаморфоз), связанное с упрощением организации. В результате развивается половозрелая фаза — материнская спороцис т а. Это очень просто устроенный организм, чаще всего мешковидной формы, способный к размножению путем партеногенеза. Партеногенетические яйца материнской спороцисты, обычно называемые генеративными или зародышевыми клетками, дробятся без оплодотворения и дают начало новому, дочернему поколению — р е д и я м. Последние, покидая материнский организм, активно ползают между органами моллюска хозяина. Обычно они обосновываются в его кровеносных лакунах (кровеносная система моллюсков не замкнута) или в пищеварительной железе. Редии, как и материнские спороцисты, размножаются только партеногенетически, отрождая либо новые поколения редий, либо своеобразных хвостатых церкарий, которые представляют собой личинок гермафродитного поколения трематод (м а р и т ы). Редии устроены сложнее, чем материнские спороцисты, так как имеют хорошо развитую пищеварительную систему, представленную мускулистой глоткой и кишечником.
Второе партеногенетическое поколение — обязательное звено в жизненном цикле трематод, но у многих высших трематод в материнских спороцистах развиваются не редии, а морфологически иные, более просто устроенные партогенетические организмы — дочерние к н о р о ц и с т ы. Об их соотношении с редиями Гтэет рассказано ниже. Здесь ограничимся яшь сведениями о том, что дочерние спороцисты лишены пищеварительной системы и имеют обычно мешковидную или шарообразную форму тела. Они редко отрождают себе подобных, как правило, давая начало лишь церкариям. Все п а р т е н и-т ы партеногенетическое поколение трематод — живородящи.
Церкарии представляют собой очень подвижных маленьких личинок, снабженных специальным органом движения — хвостом. Выйдя из тела родительской особи, они покидают организм своего хозяина-моллюска и попадают в воду. Здесь судьба их может быть различной. Церкарии одних видов довольно быстро оседают на прибрежной траве или на заглатываемых нередко птицами подводных предметах (песчинки, мелкие камешки, пустые раковинки моллюсков и т. п.). Они одеваются плотной защитной оболочкой (инцистируются) и превращаются в покоящуюся фазу, называемую адолескарией. Будучи случайно проглочена каким-либо позвоночным животным — неончательным хозяином данного вида трематод, личинка выходит из цисты (эксцистируется) и превращается в мариту. Церкарии других видов для дальнейшего развития должны попасть в организм второго промежуточного хозяина (иногда его называют дополнительным). Чаще всего это головастики, мелкие рыбки, личинки водных насекомых, моллюски, пиявки и др. В организме второго промежуточного хозяина церкарии обычно инцистируются, переходя в фазу метацеркарий. Если зараженный второй промежуточный хозяин будет съеден позвоночным животным, подходящим для развития метацеркарий данного вида, личинка выходит из цисты и достигает в окончательном хозяине половой зрелости, превращаясь в гермафродитную особь.
Таким образом, в жизненном цикле трематод обязательно участвуют три поколения — два партогенетических и одно гермафродитное. При этом происходит смена двух разных типов полового размножения: обоеполого, связанного с обязательным оплодотворением яйцеклетки, и партеногенетического, которое осуществляется без оплодотворения. В отличие от метагенеза, при котором половое размножение чередуется с бесполым. Чередование поколений, свойственное трематодам, называется гетерогонией.
Онтогенез, т.е. развитие организма от стадии яйца до половозрелости, у всех трех поколений трематод проходит различно. Онтогенез живого партеногенетического поколения — материнской спороцисты связан с метаморфозом. Ее личинкой является мирацидий, который развивается из оплодотворенного яйца мариты. Второе партеногенетическое поколение — будь то редия или дочерняя спороциста — развивается без метаморфоза. Третье, гермафродитное, поколение опять развивается с метаморфозом, но имеет не одну, а две личинки: церкарию и метацеркарию, или адолескарию.
Характерно, что первым промежуточным хозяином трематод всегда служит моллюск. Роль второго промежуточного хозяина могут выполнять самые разные животные — от кишечнополостных и гребневиков до млекопитающих включительно. Окончательный же хозяин — позвоночные. По отношению к первому промежуточному хозяину трематоды проявляют узкую специфичность. Это означает, что материнские спороцисты каждого конкретного вида трематод могут паразитировать в моллюсках лишь одного или нескольких близких видов. Напротив, ко второму промежуточному хозяину трематоды, как правило, очень широко специфичны. Метацеркарии одного вида могут поселяться в животных, относящихся не только к разным видам, но и к разным классам и даже типам. Специфичность, вернее приуроченность к тому или иному виду животных — хозяев, объясняется здесь экологическими причинами. Дело в том, что роль второго промежуточного хозяина конкретного вида трематод выполняют лишь те животные, которые служат пищей окончательному хозяину данного вида. Это важное условие для осуществления жизненного цикла паразита. По образному выражению известного немецкого ученого A. Jlooca, окончательный хозяин, выискивая свою «излюбленную» пищу, как бы сам подыскивает и собирает своих паразитов. Разумеется, из этого правила могут быть исключения: известны трематоды, которые и на стадии метацеркарий могут быть узкоспецифичны к своему хозяину.
Следует еще сказать несколько слов о взаимоотношениях между трематодами и моллюсками. Выше говорилось об узкой специфичности трематод к их первому промежуточному хозяину. Обращает на себя внимание и удивительная сбалансированность отношений между партнерами системы паразит — хозяин. Зараженный моллюск, несмотря на наличие паразитов, может долго сохранять жизнеспособность.
Морфология и физиология мариты. Обычно когда говорят о трематодах, имеют в виду паразитирующую в позвоночных животных мариту половозрелую фазу гермафродитного поколения. Ограничиться описанием только одной мариты, как это часто делается, было бы неверно. Мы уже знаем, что трематоды характеризуются сложным жизненным циклом, который включает по крайней мере три резко отличающихся друг от друга поколения. Поэтому представить особенности строения и биологии трематод можно только рассматривая каждое из поколений в отдельности. Наиболее сложно устроена марита, которая изменена паразитическим образом жизни менее других поколений и во многом сходна с организацией остальных плоских червей. С описания ее строения и биологии мы начнем наше знакомство с трематодами.
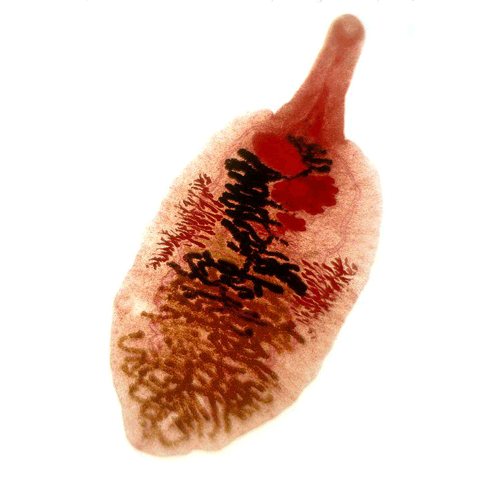
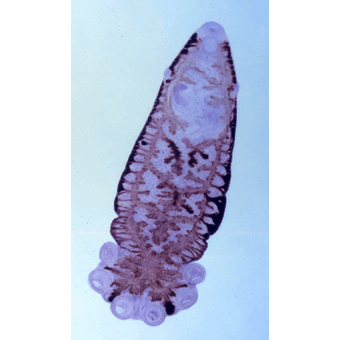
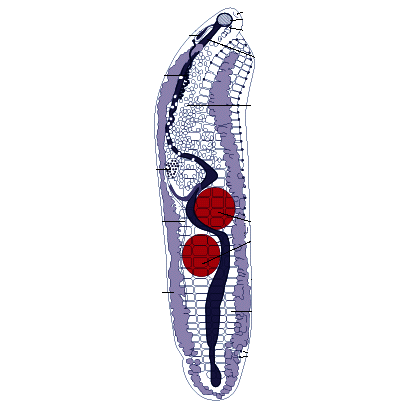
Тело мариты по большей части имеет уплощенную листовидную форму и несет одну или две присоски — органы прикрепления, снабженные сильно развитыми кольцевыми и радиальными мышцами. Передняя (ротовая) присоска всегда пронизана ротовым отверстием, вторая (брюшная) служит только органом прикрепления. Раньше считалось, что обе присоски связаны с ротовыми отверстиями. Отсюда и возникло название трематод — двуустки (Distoma).
Покровы трематод представлены типичным тегументом. Его поверхность, лишенная ресничек, часто несет переплетающиеся гребни и складки, которые играют существенную роль в питании. Слои кольцевых и продольных мышц хорошо развиты. Паренхима имеет типичное для плоских червей строение, но. в отличие от паренхимы свободноживущих турбеллярий, выполняет еще и роль депо гликогена, который служит для трематод основным источником энергии.
Пищеварительная система начинается ротовым отверстием, которое ведет в округлую мускулистую глотку, играющую роль насоса при заглатывании пищи. За глоткой следует пищевод. Он разделяется на две кишечные ветви, заканчивающиеся слепо. У крупных трематод, например у печеночной двуустки, кишечник обильно разветвлен. Эта особенность строения, как и у крупных турбеллярий, имеет важное значение в распределении продуктов пищеварения по телу червя. Длина ветвей кишечника у марит разных видов неодинакова, что связано со способом их питания. Более примитивные трематоды (Fasciola и др.) активно пожирают кровь и плотные ткани или кишечное содержимое хозяина. Иногда они «хищничают», как. например, Paramphistomum, питаясь крупными инфузориями из рубца жвачных. У таких трематод пищеварительная система развита хорошо. Напротив, у тех видов, которые лучше приспособлены к паразитическому образу жизни, большее значение приобретает способность воспринимать растворенные органические вещества через тегумент. Давно доказано, что даже Fasciola с ее хорошо развитым кишечником воспринимает через покровы глюкозу. Не удивительно, что в ходе эволюции трематод, обитающих в среде, насыщенной продуктами пищеварения хозяина, такой способ питания постепенно становится главенствующим. Одновременно с этим их пищеварительная система утрачивает значение и подвергается редукции. Нервная система устроена по типу ортогона, органы чувств развиты слабо. Выделительная система представлена протонефридиями.
Половая система трематод устроена сложно и, как у всех плоских червей, гермафродитна. Мужская половая система состоит чаще всего из двух округлых или лопастных (иногда сильно изрезанных) семенников. От них отходят семепроводы, которые, сливаясь, образуют семенной пузырь -— хранилище, в котором накапливаются зрелые сперматозоиды. Этот пузырек либо лежит в паренхиме, либо заключен в специальный мешок с мускулистыми стенками — сумку цирруса. В ней же находится и ввернутый внутрь совокупительный орган — циррус. Женская половая система представлена одним яичником, круглым или (у крупных форм) разветвленным, а также яйцеводом, желточниками, оотипом, семеприёмником, тельцем Мелиса, лауреловым каналом и маткой. В этой совокупности слаженно работающих органов собственно половой железой является яичник, в нем нормируются яйцеклетки — овоциты. Желточники продуцируют крупные клетки, цитоплазма которых забита гликогеном и содержит множество желточных или скорлуповых гранул. Благодаря им создается плотная оболочка сложного яйца. Поставляемые желточниками клетки служат для питания развивающегося зародыша. Центральная часть женской половой системы — оотип. В него открываются или из него выходят все протоки системы. Прежде всего это яйцевод, по которому в оотип поступают яйцеклетки; сюда же открывается проток маленького мешочка — семеприемника, в котором хранится семя, полученное от другой особи. По специальным протокам в оотип поступают и желточные клетки. Здесь происходит и оплодотворение яйцеклетки, и формирование сложного яйца. Овоцит окружается группой желточных клеток. Содержащиеся в них гранулы выходят наружу и окружают образовавшийся клеточный комплекс; впоследствии они дают начало плотной скорлупке сложного яйца, которое проталкивается в широкую трубку матки. В оотип открываются еще протоки одноклеточных желез тельца Мелиса, роль которых до конца не выяснена. Не исключено, что они участвуют в образовании одного из слоев скорлупки сложного яйца и выделяют секрет, играющий роль смазки при прохождении его в матку. Весь комплекс органов работает, как живой автомат, который каждые 20—30 с выталкивает в матку новое сформированное яйцо. Накапливающиеся в оотипе некоторых грематод избытки семени и желточных клеток выбрасываются наружу через короткий проток — лауреров канал, открывающийся на спинной стороне тела червя.
Матка служит для выведения яиц наружу. Но у многих видов (отряды Plagiorchiida, Cyclocoelida и др.) в яйцах, еще находящихся в матке, начинается дробление овоцита, а нередко проходит и все эмбриональное развитие личинки, так что наружу выходят яйца, в которых заключен вполне готовый мирацидий. Это очень выгодно в биологическом отношении, так как развивающиеся эмбрионы полностью избавлены от неблагоприятных воздействий внешней среды (резкие колебания температуры, высыхание и т. д.). В результате процент выживающих яиц паразита значительно возрастает. Однако эмбриогенез требует определенного времени, и развивающиеся яйца задерживаются в петлях матки, а так как поступление новых яиц непрерывно продолжается, матка становится очень длинной и ее многочисленные петли занимают иногда до 2/3 тела червя. Сами же яйца по сравнению с теми, развитие которых идет во внешней среде, становятся мельче. Это связано с тем, что развивающиеся в них зародыши получают все необходимые питательные вещества непосредственно от материнского организма. Поэтому число желточных клеток, содержащихся в сложном яйце, сокращается. Разумеется, у таких видов и сами желточники слабо развиты.
Несмотря на то, что половая система мариты гермафродитна, самооплодотворение встречается очень редко, что невыгодно для вида, так как дает генетически однородное потомство. Поэтому оплодотворение, как правило, перекрестное. Известны даже случаи появления вторичной раздельнополости марит. Таковы, например, многие представители своеобразных паразитов морских рыб из семейства Didymozoidae. Полностью раздельнополым становится все семейство Schistosomatidae — паразитов кровеносной системы человека и животных. Более крупный самец носит самку в особом «канале», который образован свернутыми в широкую трубку краями его листовидного тела.
Мариты трематод могут паразитировать в любых тканях и органах позвоночных животных. В зависимости от условий существования в тех или иных органах хозяина возникают соответствующие изменения строения и физиологии паразита. Так, мариты, поселяющиеся в кровеносных сосудах или в каких-либо узких протоках (например, желчных протоках печени), отличаются длинным, нитевидно вытянутым телом. В то же время многие тканевые паразиты имеют почти шаровидную форму. Тесно связано с местом поселения и строение органов прикрепления мариты. У паразитов кишечника, особенно у видов, обитающих в прямой кишке, откуда они легко могут быть выброшены при физиологических отправлениях хозяина, присоски развиты особенно сильно. Напротив, у трематод. паразитирующих в полости тела (например, у паразитов из семейства Cyclocoelidae), присоски дегенерируют. От локализации нередко зависит и способ питания мариты. Так, тенденция к замене активного питания всасыванием растворенных органических веществ через покровы особенно заметно проявляется у паразитов крови.
Мариты позвоночных нередко оказываются в условиях острого недостатка кислорода. Это влияет на характер обмена, который приближается к анаэробному, так как нормальное дыхание в большинстве случаев оказывается невозможным. В разных органах позвоночного животного парциальное давление кислорода может быть совершенно различным. С этим связано то обстоятельство, что трематоды не утратили способности использовать кислород, если он имеется. Поэтому им свойствен не чисто анаэробный (как предполагали раньше), а смешанный тип обмена. В зависимости от того, в каких органах поселяются мариты, в их обмене могут преобладать либо анаэробные, либо аэробные процессы. Те виды, которые паразитируют в трахее или в легких хозяина, способны к нормальному дыханию. Мариты, питающиеся кровью, по-видимому, могут использовать кислород, связанный гемоглобином крови хозяина. Наконец, в тканях многих трематод содержится гемоглобин, отличающийся повышенным сродством к кислороду. Это позволяет ма-ритам использовать даже то незначительное количество свободного кислорода, которое имеется в среде их обитания.
Трематоды, как и другие эндопаразитические черви, получают необходимую для жизнедеятельности энергию в результате расщепления глико-гена2 содержащегося в их тканях. Жиры не могут служить для них источником энергии, так как их расщепление возможно только в присутствии свободного кислорода. В тканях паразитических червей заключен» огромные запасы гликогена. У свободноживущих родичей трематод таких запасов гликогена никого не бывает. Прежде всего это обусловлено тем, что свободноживущие организмы широко используют в качестве источника энергии жиры. С другой стороны, анаэробное расщепление гликогена — процесс крайне невыгодный в энергетическом отношении. Если в присутствии кислорода гликоген расщепляется до углекислого газа и воды, то в анаэробных условиях oн распадается на крупномолекулярные соединения (главный продут-обмена — молочная кислота), npи этом высвобождается в 10 раз меньше энергии, чем в первом случае Вот почему так велики в теле мариты запасы гликогена, составляющие до 60% ее сухой массы. Не следует забывать, что много гликогена расходуется на образование бесчисленных яиц этих червей.
Первое партеногенетическое поколение
Мирацидий. В сложных яйцах, откладываемых маритой, развивается личинка материнской спороцисты — мирацидий. Развитие его может происходить во внешней среде или в матке родительской особи, но в любом случае личинка, уже полностью сформированная, может долгое время находиться внутри яйцевой скорлупки. Для выхода мирацидия из яйца требуются специальные воздействия извне: яркий свет, резкая смена температуры, недостаток кислорода или резкая смена осмотического давления. Считается, что внешние факторы стимулируют деятельность железистых клеток мирацидия, называемых железами вылупления. Секрет этих желез разрушает тонкую протеиновую полоску, соединяющую скорлупку яйца с его крышечкой. Последняя откидывается, и мирацидий выходит наружу.
В последнее время обсуждается другая точка зрения, по которой активированный внешними воздействиями мирацидий вызывает набухание заключенного в яйце комочка вязкого вещества («пробки»). В результате внутри яйцевой скорлупки создается высокое давление, под воздействием которого и происходит откидывание крышечки или разрыв скорлупки.
Мирацидий — крошечная личинка, по размерам не превышающая инфузорию. На ее расширенном переднем конце располагается короткий мускулистый хоботок. Все тело покрыто крупными плоскими клетками с бесчисленным множеством ресничек. Глубже расположена развитая система кольцевых и продольных мускульных волокон. Органов пищеварения у мирацидия нет и питаться он не может. Нервная система представлена мозговым ганглием. Из органов чувств имеются лишь пара простых пигментных глазков, сосредоточенные на переднем конце сенсиллы довольно разнообразного строения. По бокам тела симметрично расположены протонефридии с очень небольшим числом терминальных клеток (2—4). Мирацидии снабжены многочисленными железистыми образованиями, назначение которых не всегда ясно. В расширенной передней части тела располагается самая крупная, четырехъядерная железа проникновения, называемая также апикальной. Вырабатываемый ею секрет используется в процессе внедрения в тело хозяина. Протоки этой железы открываются на вершине хоботка. В задней трети тела мирацидия находится небольшая полость, в которой содержатся зародышевые клетки. Число их у разных видов колеблется от двух до нескольких десятков. Это, в сущности, партеногенетические яйца, из которых после превращения мирацидия в материнскую спороцисту развиваются особи следующего поколения.
Мирацидии большей части видов трематод — подвижные, плавающие организмы. Продолжительность их свободной жизни невелика — от нескольких часов до нескольких суток, что определяется количеством заключенного в их теле гликогена.
За короткий срок свободного существования мирацидий должен найти подходящего хозяина — моллюска и внедриться в него. Личинки, не выполнившие этой «задачи», неизбежно погибают. Существование во внешней среде требует приспособлений (адаптации), которые обеспечивали бы движение личинки и ее ориентацию. Мирацидии плавают посредством ресничек и сокращений кожно-мускульного мешка, глаза и сенсиллы помогают им ориентироваться в пространстве. Мирацидии передвигаются со значительной для их размеров скоростью, преодолевая за 1 с расстояние около 2 мм. В первый момент после вылупле-ния они плывут по прямой, словно стремясь уйти как можно дальше от места выплода. Вскоре, однако, движения их становятся зигзагообразными и они как бы «прошивают» толщу воды во всех направлениях. Эти «поисковые» движения регулируются таксисами. Личинки реагируют на световые и химические раздражения, а также на силу земного притяжения (геотаксис). Сам процесс внедрения мирацидиев в ткани моллюска изучен подробно. Важное значение имеет расположение сенсилл, сконцентрированных на переднем конце. С их помощью мирацидий находит на теле хозяина удобное для внедрения место. Секрет, нарушающий целостность покровов хозяина, по-видимому, выделяется апикальной железой. В его состав, очевидно, входят вещества, при воздействии которых покровы моллюска, в особенности коллагеновые волокна соединительной ткани, разрушаются, мускулистый хоботок механически помогает личинке внедриться в полуразрушенный участок эпителия хозяина. Этому способствуют и энергичные движения — сокращение и вытягивание тела мирацидия. Все это требует значительных затрат энергии, единственным источником которой служит гликоген. Существенно, что на протяжении всей короткой свободной жизни мирацидия гликоген в его теле как бы сберегается для решающего этапа — внедрения в моллюска. Плавание личинок в основном обеспечивается биением ресничек, которые получают энергию в результате расщепления обильного запаса гликогена в цитоплазме ресничных клеток. Гликоген тела необходим личинке не только для того, чтобы преодолеть тканевые барьеры хозяина. Попав в гемолимфу (смешанная с кровью полостная жидкость моллюска), личинка не в состоянии извлечь оттуда необходимую ей глюкозу, если запас гликогена к этому моменту истрачен. Дело в том, что транспорт глюкозы из гемолимфы в мирацидий может идти и в том случае, если в гемолимфе содержится много больше глюкозы^, чем в теле личинки (т. е. против градиента концентрации). Это активный процесс, требующий затрат энергии. Если же мирацидий, отыскивая хозяина, плавал слишком долго и истратил не только гликоген из ресничных клеток, но и заключенные в его теле «неприкосновенные запасы», то он обречен на гибель. Даже если такой мирацидий и сумеет внедриться в тело моллюска, он не сможет получить первую порцию питательных веществ, без которой невозможно дальнейшее осуществление их транспорта, а стало быть, и жизнь паразита. Поэтому сроки подвижности мирацидия всегда больше, чем сроки сохранения его инвазионности (способность заразить хозяина).
У многих трематод (например, относящихся к отряду Plagiorchiida) мирацидий вообще не выходит из яйца в воду. Он вылупляется только в кишечнике хозяина, т. е. после того как яйцо, уже содержащее сформированную личинку, будет проглочено моллюском. Отсутствие свободно плавающей стадии можно рассматривать как своеобразную адаптацию к осуществлению цикла. В самом деле, мирацидий, заключенный внутрш яйцевой скорлупки, хорошо защищен от внешних воздействий. Кроме того, он малоподвижен, поэтому его энергетические ресурсы (гликоген) расходуются очень медленно. Вследствие этого значительно увеличивается продолжительность жизни личинки. В лабораторных условиях такие мирацидии остаются живыми на протяжении 3—4 месяцев! В природе это, несомненно, повышает вероятность контакта личинки паразита с ее будущим хозяином.
Защитная роль яйцевой скорлупки позволяв мирацидиям таких видов выживать в крайне неблагоприятных условиях. Видимо, не случайно, что именно те трематоды, в жизненном цикле которых отсутствует свободноплавающий мирацидий составляют большинство среди видов, связанная в развитии с морской литоралью. В самом деле все воздействия внешней среды, которым подвигаются яйца трематод в период отлива (подсушивани, резкие колебания температуры и т. п. опасны для плавающих личинок. В еще большак степени это относится к тем видам, яйца которых попадают на поверхность земли или растениия, заглатываются наземными моллюсками. В расселении трематод, лишенных стадии свободноплавающего мирацидия, немалую роль, вероятно, играет перенос яиц дождевыми и паводковыми годами.
Материнская спороциста. Мирацидий после его проникновения в организм хозяина-моллюска превращается в половозрелую паразитически -фазу — материнскую спороцисту. Претерпеваемый мирацидием метаморфоз называется регрессивным, так как сопровождается утратой многих личиночных органов. Ресничные эпителиальные клетки обычно сбрасываются ещё во время внедрения мирацидия в тело хозяина. Тогда же расходуется и секрет апикальной железы, которая при этом разрушается. Позднее исчезают глазки мирацидия и в той или иной мере утрачиваются его разнообразные сенсиллы.
В зависимости от того, совершает ли молодая спороциста миграции в теле хозяина или нет, мускульный слой может сильно развиваться их напротив, дегенерировать. Спороцисты с развитой паренхимой встречаются редко. Обычно клетки паренхимы расходятся, образуя в центре хорошо выраженную первичную полость тела. Зрелые материнские спороцисты имеют червеобразную, мешковидную или шаровидную формы. У отдельных видов встречаются и разветвленные спороцисты. По мере роста и развития молодой материнской спороцисты начинается подготовка к выполнению ее основной жизненнее функции — размножения. Генеративные (зародышевые) клетки, которые есть уже у мирацидия, быстро увеличиваются в числе. Затем начинается дробление, которое идет без оплодотворения, т. е. партеногенетически. Эти клетки обычно не претерпевают деления созревания, соответственно партеногенез трематод диплоидный.
Внутренная организация материнских спороцист в значительной мере определяется способом их размножения. По этому признаку можно выделить две группы спороцист. К первой группе относятся те материнские спороцисты, в теле которых все зародыши (редии или дочерние спороцисты) развиваются одновременно. Как правило, у таких спороцист слабо развита паренхима. Немногочисленные паренхиматозные клетки оттеснены к стенкам тела, в центре которого образуется обширная полость. В ней и происходит развитие зародышей. Ко времени завершения их эмбриогенеза стенки материнской спороцисты сильно истончаются. Выход в полость тела моллюска уже сформированных особей дочернего поколения происходит в результате разрыва спороцисты и ее гибели. Примером спороцист такого типа может быть уже известная нам печеночная двуустка — Fasciola hepatica.
У материнских спороцист второй группы сроки развития дочернего поколения сильно растянуты во времени. Поэтому в полости материнской спороцисты одновременно находятся не только эмбрионы на самых разных стадиях развития, но и генеративные клетки. Для защиты как генеративных клеток, так и ранних стадий развития зародышей от повреждений, которые могут им нанести особи дочернего поколения, уже подвижные и готовые покинуть спороцисту, развиваются различные приспособления. По-видимому, наиболее совершенным способом изоляции разновозрастных эмбрионов следует считать так называемую терминальную массу. Это — орган, который как бы объединяет в себе гонаду и выводковую камеру. В гонаде сосредоточены интенсивно делящиеся недифференцированные клетки, дающие начало партеногенетическим яйцам. По периферии располагается выводковая камера, в которой развиваются зародыши следующего поколения. Уплощенные клетки паренхимы, окружающие терминальную массу, играют защитную роль. Эмбрионы поздних стадий отрываются и завершают развитие в полости тела спороцисты.
Возможны и другие способы изоляции разновозрастных эмбрионов. За счет остатков дегенерирующих клеток паренхимы нередко возникают особые структуры, которые образуют в полости тела спороцисты рыхлую сеть. В ячеях этой сети закреплены и как бы подвешены генеративные клетки и зародыши ранних стадий. Ко времени их созревания сеть разрушается и сформированные особи дочерних поколений через небольшие разрывы стенки тела материнской спороцисты выходят наружу. Организм ее от этого не страдает: ранка затягивается и спороциста продолжает жить и размножаться. Таким образом, со способом размножения и количеством продуцируемого потомства непосредственно связана и продолжительность жизни материнских спороцист. Виды, одновременно отрождающие незначительное число особей дочернего поколения, живут от нескольких дней до 2—3 недель. Зато материнские спороцисты с растянутым периодом размножения могут жить до 4—6 месяцев.
Второе партеиогеиетическое поколеиие
Второе партеногенетическое поколение трематод представлено двумя разными формами — редиями и дочерними спороцист а м и. Редии, как правило, свойственны примитивным семействам (Fasciolidae, Echinostoma-tidae, Paramphistomatidae и др.). Типичные редии имеют вытянутое цилиндрическое тело очень характерного облика, который придает пара локомоторных, или двигательных, выростов, расположенных в задней трети тела.
Пищеварительная система развита хорошо. Она начинается ротовым отверстием, которое ведет в мускулистую глотку; с ней связаны специальные пищеварительные железы. Длинный мешковидный кишечник почти достигает заднего конца тела. На внутренней поверхности кишечника, снабженной многочисленными микроворсинками, сосредоточены пищеварительные ферменты. Редии, особенно молодые, очень подвижны. Они активно питаются, поглощая плотные ткани и кровь хозяина. Несмотря на наличие функционирующей пищеварительной системы, типичные редии способны и к восприятию растворенных пищевых веществ извне. С этим связана и складчатость их тегумента, и концентрация на его поверхности пищеварительных ферментов. Однако, оставаясь паразитами моллюсков, редии нередко ведут себя как настоящие хищники. Они пожирают более мелких редий своего же вида или питаются спороцистами-«конку-рентами». Основываясь на этом, предполагается начать разработку метода биологической борьбы с очень опасными паразитами человека — шисто-сомами (см. «Патогенные сосальщики»). Оказывается, если моллюска, в котором развиваются спороцисты шистосом, заразить редиями вида Ribeiroia, то эти паразиты-хищники полностью уничтожают спороцист шистосом.
Выделительная система редий представлена двумя протонефридиями. Каждый из них открывается наружу самостоятельной экскреторной порой. Нервная система устроена по типу ортогона, но развита слабее, чем у мариты. Сенсиллы сосредоточены у переднего конца тела. Органом размножения служит терминальная масса, устроенная принципиально так же, как у материнской спороцисты. Сформированные особи дочернего поколения выходят наружу через специальное отверстие, называемое родильной порой, которое находится на особом бугорке позади глотки. Таким образом, дочернее поколение покидает родительский организм, не повреждая его. Редии могут отрождать дочерних редий, причем в зависимости от условий (например, зимой) отрождается несколько поколений, обильно заселяющих организм моллюска. Обычно они держатся на поверхности печени хозяина, но могут и внедряться в толщу этого органа, тканями которого питаются. Летом редии отрождают преимущественно личинок гермафродитного поколения — церкарий. Покидая материнский организм через родильную пору, эти личинки на некоторое время задерживаются в кровеносных лакунах моллюска, накапливая в своем теле необходимые запасы гликогена, без которого они не могут существовать во внешней среде. О том, каким образом эти личинки попадают в воду, будет рассказано ниже.
Строение редий разных видов трематод может довольно сильно отличаться от морфологии типичной редии. Изменения связаны главным образом с упрощением организации. Так, редии многих видов утрачивают локомоторные выросты и становятся мешковидными. Характерна и тенденция к исчезновению кишечника. Это обусловлено паразитическим образом жизни и неразрывно связано с усилением функции питания через покровы. В ходе эволюции редии именно такой способ питания приобретает главенствующую роль. Ультраструктура покровов при этом сильно изменяется, на наружной пластинке тегумента появляются многочисленные гребни и причудливые выросты, сильно увеличивающие всасывающую поверхность.
На примерах, взятых из разных групп трематод, можно построить морфологический ряд, показывающий постепенный переход от редий с длинным, хорошо развитым кишечником к таким видам, у которых кишечник представляет собой крошечный округлый мешочек, примыкающий к глотке. Есть виды (например, паразит щуки Azygia), у которых совсем нет кишечника, да и глотка видна только у молодых особей. По внешнему виду такие редии, в сущности, неотличимы от дочерних спороцист, характерная черта которых — отсутствие не только кишечника, но и глотки.
Дочерние спороцисты — другая форма второго партеногенетического поколения трематод — встречаются в основном у высших сосальщиков (отряды Plagoirchiida, Strigeidida. Schistosomatida и др.). Нередко их рассматривают как редий, которые начинают размножаться прежде, чем завершилось их развитие. Появление способности к размножению до того, как организм приобрел все черты строения взрослого животного, а иногда даже на стадии личинки, распространено широко. Особенно часто это отмечается у паразитических животных, для которых увеличение численности потомства, обусловленное ранним началом размножения, играет особо важную роль. Это явление получило название прогенеза. Но есть случаи, когда у какого-либо вида или группы близких видов прогенетическое развитие закрепляется в эволюции и животные навсегда утрачивают способность завершить развитие и приобрести облик взрослой особи. В таких случаях говорят о неотении. Есть основания полагать, что дочерние спороцисты — неотениче-ские редии. Дочерние спороцисты внешне разнообразны, они имеют мешковидную, червеобразную или шаровидную формы.
Молодые особи часто подвижны, но по мере заполнения их зародышевой полости развивающимися эмбрионами церкарий подвижность утрачивается. Многие виды поселяются, подобно редиям. в полости тела хозяина, прикрепляясь к поверхности его кишечника и других внутренних органов. Но чаще паразиты живут в гонаде или печени моллюска, причем некоторые виды (например, сем. Strigeidae) глубоко погружены в толщу печени и врастают в ее ткани.
Питаются спороцисты исключительно через покровы. Наружная безъядерная цитоплазматическая пластинка тегумента спороцисты несет очень много микроворсинок. Их строение и расположение делают поверхность тегумента очень похожей на внутреннюю стенку кишечника. Специальные железистые клетки секретируют все необхохимые пищеварительные ферменты, которые выдееляются наружу через тегумент. С помощью этих ферментов перевариваются те участки тканей хозяина, которые вплотную прилегают к телу спороцисты. Возникающие при этом первичные продукты пищеварения потом расщепляются в зоне микроворсинок до простейших аминокислот и моносахаров и всасываются через тегумент. Таким образом, на поверхности тела спороцист происходят те же процессы — пристеночное пищеварение и всасывание, что и в кишечнике других животных. Разница заключается в том, что у спороцист — паразитов, обитающих в толще -каней хозяина, весь процесс пищеварения происходит не внутри их организма, а вне его. Такой способ переваривания пищи у животных, лишенных кишечника, называется внеорганиз-м е н н ы м.
Несколько иначе питаются спороцисты из отряда Plagiorchiida, которые очень часто паразитируют не в тканях, а в гемолимфе моллюска. На их тегументе нет микроворсинок. Давно было подмечено, что стенка тела этих спороцист слагается не из одного, как обычно, а из двух слоев клеток. Но относительно недавно удалось установить истинную природу и значение наружного слоя клеток тела плагиорхоидных спороцист. Дело в том, что вокруг спороцисты, попавшей в гемо-цель моллюска, скапливаются клетки крови хозяина — амебоциты, которые оседают на поверхности ее тела. Это защитная реакция организма моллюска на появление инородного тела (каким, собственно, и является паразит). Однако паразитические животные, в данном случае спороциста, обладают удивительной способностью извращать защитные реакции хозяина, обращая их в свою пользу. Амебоциты, осевшие на тело спороцисты, вскоре принимают не свойственную им правильную цилиндрическую форму и, тесно прилегая друг к другу, одевают все тело спороцисты сплошным покровом (мантией). Одновременно тончайшие цитоплазматические выросты тегумента спороцисты внедряются между клетками мантии и, окружая их, как бы включают эти клетки в состав собственного тела. Между обоими слоями клеток образуется тесный контакт, облегчающий транспорт веществ из одного слоя клеток в другой. Клетки мантии приобретают способность извлекать из гемолимфы моллюска глюкозу, аминокислоты и другие необходимые паразиту вещества. Глюкозу клетки мантии частично передают тканям спороцисты, частично перестраивают в гликоген, который откладывается в их цитоплазме. В результате бывшие амебоциты крови моллюска превращаются в резервуары для накопления необходимого паразиту гликогена. Они становятся своего рода посредниками между паразитом и хозяином. Следует сказать о размножении спороцист. В развитии и функционировании терминальных масс здесь нет ничего принципиально нового по сравнению с тем, что было сказано в отношении других партенит. Но дочерние спороцисты, в отличие от материнских, имеют, как и редии, родильную пору, через которую выходят наружу сформированные церкарии. Продуктивность дочерних спороцист неизмеримо выше продуктивности редий. Если из моллюска, зараженного редиями, за сутки выходят во внешнюю среду сотни церкарий, то моллюск, зараженный спороцистами, за тот же срок выбрасывает в воду несколько тысяч личинок.
Личиики гермафродитного поколения
Церкарии. Церкарии представляют собой первую личиночную фазу мариты. Подобно мирацидиям, это, как правило, свободноживущие, плавающие в воде личинки. Их уплощенное листовидное тело несет на заднем конце хвостовой придаток — основной орган движения. Тело церкарии строением напоминает мариту. На переднем конце имеется ротовая, а в средней части тела — брюшная присоска. Позади последней обычно расположен зачаток половой системы, иногда более или менее развитый. Пищеварительная система по большей части вполне сформирована, но не функционирует. Церкарии, как и мирацидии, не питаются и живут за счет энергии, получаемой в результате расщепления гликогена, заключенного в теле и хвосте личинки. Нервная система развита довольно хорошо. У многих видов есть пигментные глазки. Многочисленные сенсиллы сосредоточены преимущественно на переднем конце, присосках и брюшной стороне тела. Их расположение образует правильный рисунок, постоянный в пределах систематических групп. Органы выделения (протонефридии) устроены так же, как у мариты, но отличаются значительно меньшим числом клеток с мерцательным пламенем. У церкарий имеются разнообразные по форме и функциям железы: проникновения, цистогенные (или цистообразующие),мукоидные (слизистые) и др. Наличие желез тесно связано с особенностями биологии церкарий. Те из них, которые инцистируются во внешней среде (например, церкарии печеночной двуустки), снабжены большим количеством цистогенных желез. Их секрет расходуется в момент инцистирования, образуя вокруг тела церкарий толстую защитную оболочку, которая слагается из нескольких слоев. Но желез проникновения у этих церкарий нет. Окончательный хозяин заражается случайно, проглатывая цисты.
Напротив, те виды церкарий, которые для дальнейшего развития должны сначала попасть в организм второго промежуточного хозяина, характеризуются сильным развитием желез проникновения. Их секрет помогает личинкам активно внедриться в тело хозяина. В то же время цистогенных желез у таких церкарий обычно мало, а иногда (сем. Diplostomatidae) и совсем нет. Секрет слизистых желез образует вокруг церкарии студенистую оболочку, которая защищает их во время миграции в теле моллюска или помогает укрепиться на поверхности тела того животного-хозяина, в которого они внедряются.
Церкарии, как и мирацидии, — расселительные личинки, поэтому все особенности их строения и поведения направлены на поиски тех животных, которые служат им промежуточными хозяевами, и на проникновение в них. Для этого церкарии, развивающиеся в редиях и спороцистах, прежде всего должны попасть во внешнюю среду. По большей части они выходят наружу, используя кровеносную систему моллюска. Покинув тело родительской особи, личинки проникают в вены и венозные синусы (щели между органами моллюска, по которым течет кровь), а оттуда — в сердце. Его сокращениями церкарии вместе с кровью перегоняются к органам дыхания хозяина — жабрам или же в обширные синусы мантии, через которую тоже происходит газообмен. Покровы этих органов обычно тонкие и нежные, поэтому церкарии легко пробуравливают их и попадают в воду. Наблюдая за поведением церкарий под бинокуляром, можно видеть, как на поверхности мантии образуется маленький бугорок, в котором прорывается точечное отверстие. Вскоре в нем показывается передний конец церкарии и не без труда протискивается сначала тело, а потом хвост личинки. Несколько секунд цер-кария, как бы отдыхая, еще удерживается в мантии кончиком хвоста, потом делает резкий рывок и уплывает. Церкарии других видов выходят лишь через одно отверстие, проделанное ими в мантии или жабре моллюска. Даже при небольшом увеличении хорошо видно, что идущие к этому отверстию кровеносные сосуды и синусы буквально забиты церкариями. С каждым сокращением сердца моллюска туда поступают все новые и новые «порции» личинок. Они покидают моллюска, легко выскальзывая одна за другой через пору. которая расширяется под давлением тысяч проходящих через нее крошечных тел. Церкарии — паразиты двустворчатых моллюсков приносятся кровью к жабрам, пробуравливают их и сосредоточиваются в мантийной полости. Отсюда они выносятся наружу струей воды, выбрасываемой клоакальным сифоном.
Продолжительность свободной жизни церкарии определяется тремя условиями: количеством заключенного в их тканях гликогена, температурой, которая регулирует скорость его потребления, w особенностями поведения самих церкарий. Чек холоднее вода, тем дольше живут личинки. В холодильнике при 4—5 °С их можно сохранить живыми больше недели. Но в естественных условиях, летом, когда температура воды колеблется в пределах 15—20 °С, они живут не более 1—2 дней. Если церкарии очень мелкие (например, сем. Strigeidae) и запасы гликогена в их теле незначительны, они редко выживают дольше 12 ч. Впрочем, есть виды, которые на стадии личинки могут жить довольно долго, но в таком случае они малоподвижны. Примером могут служить церкарии семейства Schistosomatidae, мариты которых паразитируют в крови млекопитающих и птиц. Поднимаясь к урезу воды, названные церкарии прикрепляются снизу к поверхностной пленке и висят неподвижно, почти не расходуя своих энергетических запасов. Они остаются живыми на протяжении 3—5 дней. В тот момент, когда рядом оказывается подходящий для них хозяин, церкарии молниеносно прикрепляются к его телу, и внедряясь через покровы, попадают в кровяное русло. Есть и другие долгоживущие церкарии, пассивно поджидающие появления второго промежуточного хозяина, роль которого выполняют разные представители морского планктона. Это личинки трематод отряда Hemiurida, заканчивающие развитие в морских рыбах. Хвост описываемых церкарий имеет причудливое строение и снабжен выростами, которые дают личинке возможность парить в толще воды. Иногда придатки хвоста расположены как спицы зонтика, которые то складываются, то раскрываются, позволяя церкарии несколько изменять свое положение. Необычен и способ заражения хозяина. Тело церкарий по большей части втянуто внутрь камеры, образованной вздутой частью хвоста. Там же помещается я особая полая внутри извергательная трубка. Когда хозяин проглатывает парящую в воде личинку, извергательная трубка выворачивается, как палец перчатки, и, действуя наподобие шприца, вводит тело личинки в ткани хозяина. Хвост со всеми его придатками отбрасывается.
В большинстве своем церкарии очень подвижные и быстро плавающие личинки. Их энергетические запасы распределены неравномерно. Дело в том, что при плавании само тело церкарии остается почти неподвижным, тогда как хвост совершает сильные гребные движения. В результате усиленной мышечной работы гликоген, содержащийся в хвосте, расходуется раньше, чем те запасы, которые сосредоточены в теле церкарии. Это имеет важное биологическое значение. Хвост представляет собой временный личиночный орган. Как только церкария прикрепляется к телу хозяина, он отбрасывается. Сама же личинка начинает вбу-равливаться в ткани, что сопровождается энергичными движениями ее тела, требующими значительных затрат энергии. Но и после внедрения личинке необходима энергия, чтобы обеспечить получение первых порций питательных веществ.
Поиски хозяина, необходимого для дальнейшего развития церкарии, далеко не всегда завершаются успешно. По мере того как расходуется гликоген, сосредоточенный в хвосте, движения личинки становятся все более вялыми. Она теряет способность плавать и опускается на дно, где живет еще некоторое время за счет потребления гликогена тела. Каким же образом отыскивают церкарии необходимого хозяина? В их поведении можно заметить много общего с поведением мирацидиев. Но в выборе хозяина церкарии, пожалуй, менее разборчивы. Успех поисков обеспечивается сочетанием таксисов, работы сенсилл, способа передвижения и даже характером суточного выхода церкарий из моллюска в воду. В отличие от мирацидиев, церкарии обычно появляются в воде в массовых количествах. Иногда церкарии выходят из моллюска равномерно, на протяжении всего светлого времени суток. Обычно это свойственно тем трематодам (например, печеночной двуустке), жизненный цикл которых проходит без участи второго промежуточного хозяина. Покинув моллюска, церкарии названного вида поднимаются к урезу воды. Личинки оседают на прибрежной траве или на поверхностной пленке воды. Они отбрасывают хвост и инцистируются, одеваясь плотной прозрачной оболочкой. В таком состояния личинки (теперь они называются адолескариямя) могут оставаться живыми очень долго, даже в скотонном и высушенном сене. Овцы и коровы заглаживают их вместе с травой, водой, сеном и заражаются. У тех трематод, жизненный цикл которых связан с наличием второго промежуточного хозяина, церкарии выходят из моллюска лишь в строго определенное время суток. Если хозяин ведет ночной образ жизни, то и массовый выход циркарий обычно приурочен к ночному времени; если он активен днем, церкарии, как правило, выедят днем. Немалое значение имеет и температура, стимулирующая массовый выход церкарий. Поясним роль этих адаптаций на конкретном примере.
Церкарии сосальщика Maritrema subdolum, который во взрослом состоянии паразитирует в кишечнике птиц, развиваются в спороцистах — паразитах морских брюхоногих моллюсков (Hydrobia ulvae). В условиях эксперимента церкарии этого вида выходят один раз в сутки и только при температуре 28—30 °С. Между тем Н. ulvae — Читатели северных морей, где такая температура годы обычно не отмечается. Однако во время отлива на обсыхающей литорали, между камнями или в неровностях дна, всегда остаются небольшие лужицы. На Белом море в теплый солнечный день вода в таких лужицах быстро прогревается до 25—29 °С. Сюда и сползаются моллюски, различные ракообразные и другие мелкие беспозвоночные. Повышение температуры служит сигналом для массового выхода в воду церкарий М. subdolum, но эти личинки, выходящие из моллюска днем и при ярком солнце, проявляют отрицательный фототаксис и тотчас же устремляются под прикрытие водорослей (фукусов) или мелких камешков. Это непонятное на первый взгляд явление объясняется просто. Вторым промежуточным хозяином М. subdolum служат бокоплавы Gammarus locusta, которым свойствен отрицательный фототаксис. В лужицах эти рачки всегда держатся под камнями или фукусами, и церкарии благодаря особенностям их поведения попадают именно туда, где их встреча с хозяином наиболее вероятна. Когда церкарии оказываются в зоне обитания хозяина, возможность войти в соприкосновение с ним становится реальной. Но роль таксисов на этом не кончается. Если хозяин ведет придонный образ жизни, церкарии, направляемые положительным геотаксисом, как правило, опускаются на дно. Если хозяин живет в поверхностных слоях воды, геотаксис у церкарий обычно отрицательный. Все это удается наблюдать в экспериментальном аквариуме. Но в природе, где среда обитания неизмеримо более разнообразна, действие таксисов проявляется не столь прямолинейно. Вопрос о том, как ведут себя церкарии в естественном водоеме, нуждается в специальном изучении.
Большая роль принадлежит и хемотаксису. Реакции церкарий на исходящие от хозяина химические воздействия представляют собой впечатляющее зрелище. При изучении жизненного цикла одного вида вилохвостых церкарий в солонку с густой взвесью активно плававших личинок были помещены самые разные животные. Церкарии не реагировали на их появление, но когда в аквариум попала пиявка (Herpobdella octoculata), поведение церкарий резко изменилось. Через несколько минут пиявка была буквально облеплена сотнями личинок, которые ползали по ее телу и активно внедрялись в покровы.
Сам процесс внедрения происходит в несколько этапов. Прежде всего церкария должна хорошо закрепиться на поверхности тела хозяина, который резкими движениями пытается сбросить ее. Удержаться на теле хозяина церкарии помогают ее мускулистые присоски и клейкий секрет слизистых желез. Личинка быстро отбрасывает хвост и ползает по телу хозяина, отыскивая подходящее для внедрения место. Вероятно, важную роль при этом играют сенсиллы, сосредоточенные на ее переднем конце. «Определив» посредством сенсилл подходящее место, церкария располагается перпендикулярно к поверхности тела хозяина. С этого момента начинается второй этап — повреждение покровов хозяина. Многие церкарии (отряд Plagiorchiida и др.) снабжены сильным кутикулярным стилетом, имеющим вид копья или обоюдоострого ножа, который расположен на переднем конце тела и приводится в движение специальными мускулами. Стилетом церкария надрезает хитин членистоногих или кожу позвоночных животных. У других церкарий (отряд Strigeidida, Schistosomatida) развит особый «передний орган». Он располагается на место ротовой присоски и представляет собой короткий и сильный хоботок, усаженный рядами крючьев, острия которых загнуты назад. Передний орган, подобно пальцу перчатки, может вворачиваться и вновь выворачиваться, царапая крючьями поверхность кожи хозяина. В нанесенную ранку церкария вводит секрет желез проникновения, протоки которых открываются на самом переднем конце ее тела. Этим секретом являются ферменты, переваривающие белки, протеазы, гиалуронидаза, которая разрушает межклеточное вещество, и др. Последний этап заражения хозяина сводится к тому, что церкария энергичными мускульными усилиями (попеременное сокращение и вытягивание тела) постепенно внедряется в толщу поврежденного ею участка покровов. Внедрившаяся личинка паразита вскоре инцистируется в толще тканей или во внутренних органах хозяина, окружаясь плотной, но тонкой оболочкой. С этого момента она называется метацеркарией (вторая личиночная фаза мариты).
Далеко не все церкарии способны к активному внедрению через покровы. Многие виды попадают в хозяина пассивно. Особенно любопытны адаптации к пассивному заражению, связанные со своеобразной мимикрией. Явление мимикрии широко распространено среди насекомых. Суть его сводится к тому, что незащищенные виды подражают своим обликом и окраской защищенным насекомым, ядовитым или жалящим. Церкарии и спороцисты напоминают внешним обликом мелких животных, которыми обычно питаются рыбы или птицы. Встречается это явление не так уж часто. Классическим примером могут служить церкарии рода Azygia, имеющие огромный по сравнению с телом хвост, длина которого достигает нескольких миллиметров. Хвост окрашен в темный серо-коричневый цвет и, непрерывно изгибаясь, напоминает движением, цветом и общим обликом личинок комаров. Мелкие рыбки с жадностью набрасываются на церкарий и поедают их. Огромный хвост переваривается, а сама церкария прикрепляется к стенке желудка. Она длительное время остается живой, но не растет. Только в том случае, когда зараженная рыбка становится жертвой щуки, паразит приживается в желудке хищника и достигает там половозрелости. Мелкие рыбки, питающиеся планктоном, играют в этом цикле роль резорвуарного хозяина, в котором происходит накопление паразитов, причем последние живут, но не растут и не развиваются.
Огромными, окрашенными в темный цвет хвостами, не образующими камер на переднем конце, обладают и некоторые церкарии семейства Ecfcnostomatidae. Чаще всего они заканчивают развитие в птицах, а рыбы, проглотившие церкари играют роль второго промежуточного хозяна. Особенно интересны некоторые представители рода Echinochasmus, церкарии которых соединяются кончиками хвостов и образуют крупные, хорошо заметные в воде плавающие шары (рис. Они напоминают шаровидные колонии коловраток и охотно поедаются рыбами. Когда шар проглочен, церкарии молниеносно отрываются от объединенных хвостов и инцистируются на жабpax рыбы.
Наиболее яркий пример мимикрии дает жизненный цикл сосальщиков из рода Leucochloridium — паразитов птиц. Если пройти вдоль берега небогатого ручья, реки, пруда или озера, то на прибрежной растительности, часто у самой воды можно увидеть множество небольших наземных улиток—янтарок, получивших название за цвет их прозрачной раковины. Внешне все улитки глядят одинаково, но при беглом осмотре показаться, что впереди одной или двух улиток ползут маленькие, ярко окрашенные полосатые гусеницы, чаще всего зеленые или коричневые. Присмотревшись к ним внимательнее, можно видеть, что гусеницы не ползут, они как будто приклеены к улитке, и только непрерывные сокращения их тела создают иллюзию движения. На самом деле это вовсе не гусеницы, а лишь похожие на них спороцисты сосальщиков Leucochloridium, которые заключены внутри полых и совершенно прозрачных щупалец моллюска. Впервые эти удивительные спороцисты были обнаружены еще в начале прошлого века немецким зоологом К а р у с о м (1835), который сам был поражен сделанным им открытием. Вскрыв спороцист и обнаружив в них неполовозрелых трематод, ученый определил систематическое положение этих животных, которых назвал Leucochloridium paradoxum (в переводе «бело-зеленый удивительный»). Однако жизненный цикл названного вида удалось раскрыть лишь много десятилетий спустя.
Улитки заражаются, проглатывая попавшие на лпстья растений яйца сосальщиков. В яйцах уже содержатся мирацидии, которые, вылупляясь и кишечнике моллюска, внедряются в его печень и вскоре превращаются в своеобразную разветвленную спороцисту. Развивающиеся в ней церкарии не имеют хвоста и никогда не выходят во внешнюю среду. Ветви спороцисты растут неравномерно. Самые крупные из них, заполненные развитыми церкариями, приобретают подвижность. Не отрываясь от общего столона, они пробираются между органами хозяина и проникают в его щупальца, непомерно растягивая их. Толстые мешковидные ветви спороцисты приобретают яркую окраску, чаще всего зеленую или бурую, нередко с белыми полосами и темными красно-коричневыми пятнами. Окраска и пульсирующие движения спороцист привлекают внимание птиц. Это можно доказать простым опытом.
Если в большую кювету поместить сотни янтарок и среди них одну со спороцистами, пущенный туда птенец, способный сам хватать пищу, немедленно склевывает единственного зараженного моллюска, не обращая на остальных внимания. В кишечнике птицы церкарии высвобождаются из спороцисты, которая переваривается, и прикрепляются мощными присосками к стенке прямой кишки, где и превращаются в мариту.
Мы привыкли рассматривать трематод как паразитов, вызывающих тяжелые заболевания, причиняющих большой хозяйственный ущерб, а иногда угрожающих здоровью и даже жизни человека. Это верно. Но чем глубже становятся наши знания о взаимоотношениях между животными в пределах отдельных экологических систем, тем чаще убеждаемся в том, что безусловно вредных животных не бывает, что все они играют ту илп иную (подчас очень важную) роль с природе. Исследования последних лет, проводившиеся под руководством московских гельминтологов А. А. Шпгпна и В. Е. Судариков а, показали, что трематоды не составляют исключения ну этого общего правила. Церкарии, а возможно и мирацидии, составляют существенную часть планктонных сообществ. Эти личинки, тело которых содержит очень высокий процент гликогена, несомненно, представляют собой полноценную пищу для мелких планктонных животных.
Церкариями питаются многие коловратки. Эти крошечные планктонные хищники заглатывают мелких церкарий целиком. Но они способны нападать и на добычу, превышающую их по размерам, и, разрывая тело жертвы, жевательным аппаратом пожирают ее. Одна коловратка за свою недолгую жизнь съедает около 400 церкарий. Коловратки нормально растут и размножаются даже в том случае, если их кормят одними церкариями. Другой потребитель личинок трематод — малощетинковый червь хетогастпер (Chaetogaster limnaei), живущий в мантийной полости многих пресноводных улиток. Он относится к истинным комменсалам и питается не за счет своего хозяина, а мелкими планктонными животными. Тело хетогастера совершенно прозрачно; рассматривая его под микроскопом, можно легко определить состав поглощаемой им пищи, которая просвечивает через стенки кишечника. Помимо мелких коловраток, простейших и одноклеточных водорослей, там всегда можно обнаружить мира-цидиев и церкарий. Удается длительное время содержать хетогастеров на «диете», состоящей из одних церкарий. Червп при этом не только живут, но и интенсивно размножаются. Активно пожирают церкарий и планктонные рачки — дафнии п циклопы. Расчеты показывают, что в природных очагах диплостоматоза (тяжелого заболевания рыб, вызываемого церкариями рода Diplostomun) рачки выполняют роль санитаров,, так как выедают практически всех выходящих в воду церкарий. Питаются ими и многие водные насекомые, например клопы Corixa, личинки стрекоз и поденок. Наконец, церкарий охотно едят и мальки рыб. Показательны в этом отношении результаты опыта, в котором два малька воблы, содержавшиеся в аквариуме с зараженными моллюсками, уничтожали за сутки около 4—5 тыс. личинок. На первый взгляд питание мальков церкариями кажется парадоксальным — ведь многие из них паразитируют в рыбах и вызывают тяжелые заболевания. Но не все церкарии используют рыб в качестве хозяина, а те из них, которые становятся паразитами, проникнув через покровы рыбы, будучи проглочены, не приживаются, а перевариваются. Все это заставляет по-новому взглянуть на роль трематод в биоценозах. Борьбу с трематодами, опасными для человека и животных, следует осуществлять на строго научной основе, помня о том, что за миллионы лет эволюции экологических систем они настолько сбалансированы, что грубое и необдуманное вмешательство человека может привести к нежелательным, а порой и непредвиденным результатам.
Метацеркария. Следующая за церкарией фаза личиночного развития мариты носит название метацеркарии, если личинка инцистиру-ется в организме второго промежуточного хозяина, или адолескарии, если инцистирова-ние происходит во внешней среде. По строению и биологии личинки этих двух типов несколько отличаются друг от друга.
Адолескария одета плотной защитной оболочкой, которая состоит из 2—4 слоев, предохраняющих личинку от неблагоприятных воздействий внешней среды. Свернувшаяся в цисте адолескария почти неподвижна, и поэтому запасы гликогена в ее паренхиме расходуются очень медленно. Это позволяет личинке сохранять жизнеспособность в течение длительного времени, но она не растет и не претерпевает никакого развития. Метацеркарии, паразитирующие во втором промежуточном хозяине, как правило, заключены в цисты с очень тонкой, по большей части однослойной стенкой. Иногда они вообще не инцистируются (например, сем. Diplostomatidae, Gymnophallidae) и лежат в тканях хозяина свободно или даже передвигаются в них. Личинки питаются за счет хозяина и растут, иногда значительно увеличиваясь в размерах. Нередко на этой фазе развития начинается (или полностью завершается) развитие органов половой системы. В таких случаях (например, у сосальщиков сем. Microphallidae) паразиты обнаруживают узкую специфичность по отношению к хозяину. Иногда метацеркарии даже начинают откладывать яйца, которые накапливаются внутри цисты, но, как правило, не попадают наружу. Ведь обычно метацеркарии локализуются в мышцах, в полости тела или во внутренних органах хозяина, изолированных от внешней среды. Впрочем, у многих видов связь метацеркарий с внешней средой сохраняется и обусловлена способом заражения хозяина. Так, личинки некоторых представителей семейства Echinostomatidae (Petasiger и др.) засасываются рыбой вместе с водой и могут инцистироваться на стенках ее ротовой полости или на постоянно омываемых водой жабрах. Личинки других эхиностоматид, оседая на поверхности тела улиток, заползают в их дыхательное отверстие и инцистируются на внутренних стенках легочной полости.
Для многих трематод характерно активное внедрение в покровы рыб, амфибий или водоплавающих птиц. Метацеркарии инцистируются непосредственно в месте внедрения — в мышцах или подкожной клетчатке. Но немало и таких видов, которые, внедрившись в хозяина, длительно мигрируют по его телу, используя кровеносные сосуды или передвигаясь прямо через ткани. Они двигаются до тех пор, пока не достигнут места своего окончательного поселения — очень часто в мозге или в глазах рыбы. Это «путешествие» сопровождается многочисленными внутренними кровоизлияниями, в результате которых наступает серьезное заболевание, а иногда и гибель хозяина, особенно при массовом нападении церкарий. Больные рыбы делаются вялыми, часто держатся ближе к поверхности водоема, где становятся легкой добычей чаек, в кишечнике которых паразиты вырастают до взрослого состояния.
Молодые мариты, т. е. метацеркарии, только что проглоченные окончательным хозяином, тоже часто проделывают в его теле длительные миграции. Дело в том, что паразит, попав в кишечник хозяина, как правило, стремится покинуть его (так называемая «реакция бегства»), потому что на этой стадии развития он еще не способен противостоять ни воздействию пищеварительных ферментов, ни перистальтическим сокращениям кишечника, выносящим его наружу. Паразиты проникают в полость тела или кровеносные сосуды и. проделав миграцию, либо возвращаются в кишечник, претерпев физиологические и морфологические изменения, необходимые для жизни в этом органе, либо поселяются в других местах. Так. проглоченные с травой адолескарии печеночной двуустки, высвободившись из цисты, проходят сквозь стенку кишки млекопитающего и оказываются в полости его тела. Они пробираются к печени, внедряются в нее и начинают поглощать клетки печеночной паренхимы. Паразиты в полном смысле слова «выедают» себе дорогу, пробираясь через толщу печени к желчным протокам, в которых и поселяются. Подобные, а порой и гораздо более сложные миграции описаны и для многих других трематод (Paramphistomum, Cyclocoelum, Alaria и др.). Нередко именно в этот период жизни паразиты особенно опасны для хозяина.
ТРЕМАТОДЫ — ВОЗБУДИТЕЛИ ЗАБОЛЕВАНИИ ЖИВОТНЫХ И ЧЕЛОВЕКА
Среди трематод известно немало видов, которые, поселяясь в тех или иных органах человека и животных, могут вызывать тяжелые нарушения в их работе и становятся причиной серьезных, иногда даже смертельных болезней.
Фасциолез. Печеночная двуустка, о которой не раз шла речь в этой книге, паразитирует в желчных ходах печени мелкого и крупного рогатого скота, в редких случаях встречается и у человека. Это опасный паразит, жизненный цикл которого проходит с участием лишь двух хозяев. Первым служит пресноводная улитка малый прудовик (Lymnaea truncatula), вторым (окончательным) — различные млекопитающие и человек. Мариты отличаются очень высокой плодовитостью. Яйца для развития непременно должны попасть в воду. При благоприятных температурных условиях через 17—18 дней из них выходят мирацидии, которые, поднимаясь к урезу воды, внедряются в мантию или ногу улитки, проникают в кровеносные лакуны и превращаются в мешковидные материнские спороцисты. В последних развиваются редии, которые, вырастая, разрывают спороцисту, попадают в полость тела моллюска и внедряются в его печень. Через 30—70 дней от начала заражения в воду начинают выходить церкарии. Они поднимаются к урезу воды и инцистируются на траве или на поверхностной пленке. Скот заражается, проглатывая адолескарий с травой или водой, человек — при питье сырой воды из мелких водоемов, часто образующихся на заболоченных лугах. Именно в таких временных водоемах охотно поселяются малые прудовики. Поэтому в дождливые годы фасциолез распространен шире, чем в засушливые. Это тяжелое заболевание особенно опасно в то время, когда личинки паразита мигрируют через печень, разрушая ее. Больные животные становятся вялыми, худеют, у коров снижаются надои молока. Но особенно опасен фасциолез для молодняка. Сейчас благодаря хорошо разработанным приемам лечения и профилактическим мероприятиям (осушение заболоченных пастбищ, уничтожение моллюсков и т. д.) распространение фасциолеза сократилось.
Фасциолез у человека протекает в тяжелой форме, в остром периоде связанной с высокой температурой, сильными болями, желтухой, расстройствами пищеварения и другими тяжелыми симптомами. Чтобы уберечься от фасциолеза, нужно помнить о том, что нельзя пить воду даже, казалось бы, из чистых луж на заболоченных пастбищах.
Дикроцелиоз — заболевание печени рогатого скота (преимущественно овец), связанное с поселением в желчных протоках другого сосальщика — ланцетовидной двуустки Dicrocoelium dendriticum (или D. lanceolatum).
Болезнь протекает сходно с фасциолезом, но в смягченной форме. Заболевание животных сказывается на их продуктивности. Гибель отмечается не часто, хотя степень заражения может быть очень высокой — до 50 тыс. червей у одного животного. Заражение человека хотя и возможно, но встречается редко, что связано с особенностями крайне своеобразного жизненного цикла ланцетовидной двуустки.
На протяжении долгого времени жизненный цикл этого сосальщика оставался загадкой. Лишь в 30-х годах нашего столетия известный немецкий зоолог Н е й г а у з опубликовал результаты своих работ, раскрывших ход развития ланцетовидной двуустки. Оказалось, что ее первым промежуточным хозяином служат не пресноводные, а наземные улитки из родов Helicella, Zebrina, Theba и др. Откладываемые маритой зрелые яйца уже содержат в себе мирацидиев, которые не выходят во внешнюю среду и потому не имеют глаз. Яйца, выведенные из кишечника хозяина, попадают на траву или поверхность земли. Они обладают очень плотной и сложно устроенной скорлупкой, благодаря чему выдерживают высыхание, замораживание и воздействие некоторых химических соединений. Если улитка проглотит такое яйцо, то в ее кишечнике крышечка яйца открывается, мирацидий выходит из скорлупки и внедряется в печень хозяина, превращаясь в разветвленную материнскую спороцисту. В ней развиваются дочернне спороцисты, которые начинают отрождать церкарий. Это происходит через 4—5 месяцев после того, как улитка проглотила яйцо. Мы уже знаем, что церкарии — в большинстве случаев свободноплавающие личинки. Но как же смогли приспособиться к условиям существования церкарии, развившиеся в наземной улитке? Оказалось, что, покинув спороцисту, они проникают в легочную полость моллюска и накапливаются там. Выделяя обильную слизь, личинки слипаются в большие комки. При дыхательных движениях моллюска эти цисты при каждом выдохе одна за другой выдавливаются через легочное отверстие наружу. За ползущей улиткой на траве остаются светлые слизистые комочки. Наружный слой слизи на воздухе быстро подсыхает, и под образовавшейся тонкой плотной пленочкой церкарии остаются живыми.
Собрав травинки с прилипшими к ним слизистыми комочками, ученый скормил их ягненку. Через некоторое время в печени ягненка было обнаружено множество ланцетовидных двуусток. Открытие Нейгауза стало сенсационным, многолетняя загадка была разрешена.
В начале 40-х годов дикроцелиоз широко распространился в США. Американский паразитолог К р а л л решил повторить опыты Нейгауза. Однако сколько бы он ни скармливал ягнятаи сборной цисты, заразить животных ланцетовидной двуусткой не удавалось. Оказалось, что в жизненном цикле ланцетовидной двуустки имеется еще один, второй, промежуточный хозяин. Не овцы, а рабочие муравьи-фуражиры, собирающие пищу для обитателей муравейника, подхватывают с травинок слизистые комочки. В кишечнике муравья, проглотившего сборную цисту,церкарии освобождаются от слизи и выходят черег стенку его кишечника в полость тела. Здесь они инцистируются, становясь метацеркариями. Но одна личинка сохраняет подвижность. Пробираясь между внутренними органами муравья, он продвигается к голове и в конце концов внедряется в мозговой ганглий, где и остается, неинцисти-руясь. Постоянное присутствие паразита в мозг* насекомого оказывает сильное воздействие на его инстинкты и поведение. В течение дня муравей ведет себя как обычный фуражир. Но к вечер>. когда его собратья уходят в гнездо, зараженныж фуражир всползает на травинку, крепко сжимает ее челюстями и, надолго потеряв способность е движению, повисает на травинке до утра. В очагам дикроцелиоза возле муравейников всегда мож&. видеть этих неподвижно висящих муравьев. Их вместе с травой и поедают овцы, заражаясь таким образом метацеркариями ланцетовидной двуустки.
В чем же заключалась ошибка Нейгауза, первым исследовавшего жизненный цикл Dicrocotlium dendriticum? Вероятно, служитель, ухаживавший за подопытными овцами, не доглядел за ягненком, получившим порцию сборных цист Видимо, животное успело съесть пучок травы висящим на ней муравьем и заразилось метацеркариями, что исказило результаты опыта.
Описторхоз. Заболевание вызывается кошачьей или сибирской, двуусткой Opisthorchis felines, мариты которой тоже паразитиружг в желчных ходах печени млекопитающих. Как и у ланцетовидной двуустки, в зрелых яйцах япогх паразита заключены уже вполне развитые мирацидии, лишенные глазка. Яйца должны попасть в воду, но мирацидии из них не выходят до тех пор, пока не будут съедены первым промежуточным хозяином — моллюском из рода Bithynia. Эгх небольшие улитки с крепкой башневидной рак-винкой, устье которой животное может закрывать известковой крышечкой. Дышат они не легкюа а жабрами и относятся к так называемым переднежаберным моллюскам (Prosobranchia). Из пр:-глоченных битинией яиц выходят мирацидкж Пройдя сквозь стенку кишки д они оказывает в гемоцеле моллюска и там превращаются в материнские спороцисты, в которых развиваются редпи. Размножение и развитие партенит О. felineus продолжается очень долго — до полугода. Только по истечении этого срока в воду начинают выходить крупные, подвижные церкарии, длинный хвост которых снабжен хорошо развитой плавательной мембраной.
Церкарии оседают на тело многих видов пресноводных рыб (плотва, линь, лещ и др.) и, проникнув через покровы, инцистируются в мускулатуре. Окончательным хозяином служат различные рыбоядные млекопитающие, в том числе и домашние кошки, которых кормят рыбой. Очень часто заражается этим сосальщиком и человек. Хотя природный очаг описторхоза довольно широк, заражение человека ограничено в основном Сибирью, где принято употреблять в пищу мороженую рыбу или очень слабо соленую. При таком приготовлении метацеркарии остаются живыми и человек получает большую дозу паразитов, которые вызывают тяжелое и опасное заболевание. Хотя болезнь встречается и в европейской части нашей страны, главный центр ее распространения — бассейны рек Оби и Иртыша.
Болезнь протекает тяжело, с нарушениями в работе печени (воспаление желчного пузыря, желтуха). Чтобы избежать заболевания, необходимо употреблять только тщательно обработанную рыбу. Метацеркарий убивает горячее копчение, любая термическая обработка и засолка продолжительностью не менее 3—4 суток. К замораживанию паразиты довольно устойчивы, и крупную рыбу нужно держать при температуре —12 °С не менее 2—3 недель, чтобы рыба стала безопасной.
Шистосоматозы. Под этим названием объединяют заболевания, вызываемые тремя видами сосальщиков из рода Schistosoma: Sch. mansoni, Sch. haematobium и Sch. japonicum. Первые два вида — специфичные паразиты человека, третий встречается, кроме того, и у некоторых домашних животных. Все шистосомы паразитируют в крови хозяина.
Шистосоматоз — болезнь южных и тропических стран, широко распространенная в Африке, Юго-Восточной Азии и Южной Америке. По данным Всемирной организации здравоохранения, в мире насчитывается* около 200 млн. человек, страдающих шистосоматозом. Вот почему над изучением биологии и физиологии шистосом, их жизненного цикла и географического распространения, симптомов болезни, способов ее лечения и предотвращения работает целая армия биологов и врачей.
Жизненный цикл у разных видов шистосом протекает одинаково и изучен достаточно хорошо. В нем участвуют два животных-хозяи-на: первый — моллюски, второй — человек, домашние и дикие млекопитающие, а для некоторых видов и водоплавающие птицы. Половозролые черви обычно поселяются в венозных сосудах. Sch. mansoni и Sch. japonicum — возбудители так называемого кишечного шистосоматоза — локализуются в мезентериальных венах. Sch. haematobium, вызывающий поражения мочевого пузыря и почек, обитает в венозных сплетениях мочеполовой системы. Взрослая самка шистосомы откладывает каждые 7—8 мин по одному яйцу непосредственно в кровь, где и происходит развитие мирацидия. С током крови яйца разносятся по телу больного и в конце концов задерживаются в капиллярах. Это приводит к их закупорке, разрывам и разрушению, что вызывает образование тромбов и мелких кровоизлияний. В результате яйца оказываются лежащими прямо в тканях и становятся центрами воспалительных очагов и абсцессов (нарывов), которые могут прорваться в кишечник или в просвет мочевого пузыря. Отсюда яйца выносятся во внешнюю среду. Развитие мирацидиев к этому времени обычно уже завершено, и как только яйцо попадает в воду, скорлупка его лопается (яйца шистосом не имеют откидывающейся крышечки) и мирацидий высвобождается. Происходит это в результате резкой смены осмотического давления.
Первым промежуточным хозяином шистосом служат ярко-красные улитки-катушки рода Biomphalaria, хорошо известные любителям аквариумов, улитки Bulinus с башневидными раковинками и переднежаберные (с крышечками) улитки Oncomelania. В теле моллюска мирацидий превращается в материнскую спороцисту, которая отрождает множество дочерних. Последние очень плодовиты: из моллюска, зараженного только одним мирацидием, ежедневно выходит в воду около 3500 церкарий, относящихся к одному полу. В крови окончательного хозяина потомки одного мирацидия могут давать начало либо только женским, либо только мужским особям.
Церкарии шистосом очень мелкие, размер их тела не превышает 0,2 мм. Хвост вилообразно расщеплен на конце и очень подвижен. С его помощью церкарииж как штопор, ввинчиваются в водную толщу, а поднявшись, неподвижно провисают, прикрепляясь брюшной- присоской поверхностной пленке. При соприкосновении с телом хозяина — купающегося или работающего в воде человека — они тотчас же нападают на него, с трудом преодолевая толстый роговой слой человеческой кожи. Через живую точную часть покровов церкарии проходят очень быстро. Весь процесс их внедрения тщательно изучен и занимает около 15 мин. Попав в кроивяное русло, личинки шистосом сначала заносятся ком крови в легкие, потом в печень и наконец на 20—30-й день в вены.
Уже сам процесс внедрения церкарий сопровождается сильным зудом, покраснением кожи, крапивницей. Миграция паразитов в легкие вает кашель и кратковременное повышение температуры. Следующие 2—3 месяца человек казался здоровым, но потом наступает острый период болезни, характеризующийся высокой температурой, ознобами, сильной слабостью и исхуданием. После кратковременного периода кажущеся выздоровления начинают проявляться результаты повреждения пораженных паразитами органов: печени, кишечника, почек и мочевого пузыря. Заболевание сопровождается многочисленными побочными явлениями: нарушениями сердечной деятельности, образованием абсцессов в местах скопления яиц, разрастанием тканей, которое приводят к возникновению различных опухолей. Болезнь принимает хронический характер и может длиться годами. Болеют и взрослые, и дети. 3аражение возможно только в воде, и поэтому оно либо связано с купаньем, либо носит профессиональди характер. Чаще всего болеют крестьяне, возделывающие залитые водой рисовые поля, рабочие, возводящие мосты, дамбы, и т. д. Лечить шистоматоз трудно.
В нашей стране шистосоматоза нет, но распространено заболевание, известное под названием «зуд купальщиков». Оно связано с тем, что цистерии шистосом, паразитирующих в птицах harzia и др.), могут проникать в кожу человека-, вызывая болезнь, соответствующую первой стадии истинного шистосоматоза. Птичьи паразиты при сильных заражениях могут спровоцировать и вторую (легочную) стадию, но на этом все и кончается, так как шистосомы птиц не могут существовать и развиваться в организме неспецефичного для них хозяина — человека.
Трематодозы рыб и птиц. Выше уже говорилось том, что животные, которые служат для трематод вторым промежуточным хозяином, особенно сильно подвержены болезнетворному воздействию паразитов. Так, тяжелые и даже смертельные заболевания рыб, особенно молоди, вызывают метацергирии сосальщиков из семейства Diplostomatidа: заканчивающие развитие в кишечнике рыболовных птиц. Зараженные рыбы болеют и гибнут в результате кровоизлияний, возникающих по пути миграции метацеркарий. Поселение их в хрусталике глаза рыб приводит к слепоте (паразитарная катаракта). Губительна и характерная для некоторых видов локализация метацеркарий в мозге рыб.
Известны случаи массовой гибели (эпизоотии) рыб и в вырастных прудах рыбоводных хозяйств, И в природных условиях. Обычно это происходит вблизи от колоний чаек, рассеивающих яйца сосальщиков названного семейства.
В дельтах больших рек — Волги, Днепра, Дуная и др. нередко можно видеть рыб, поверхность тела которых испещрена черными пятнами. Это заболевание, известное под названием черно-пятнистой болезни, очень опасно, особенно для мальков. Вызывается оно метацеркариями сосалыщикa Posthodiplostomum cuticola. Первым промежуточным хозяином данного паразита служат улитки-катушки (Planorbis), а окончательным — цапли, которые обычно селятся колониями и распространяют очень большое количество яиц. Метацеркарии P. cuticola инцистируются в подкожной клетчатке рыб. Вокруг каждой цисты вскоре обязуется толстая соединительнотканая капсула, в которой накапливается черный пигмент, — так возникают пятна, придающие больной рыбе характерный облик. У зараженных рыб резко снижается количество гемоглобина, изменяется формула крови, они сильно худеют и погибают.
Из опасных паразитов диких птиц отметим лишь сосальщика Microphallus pygmaeus — возбудителя тяжелого заболевания ценной промысловой птицы гаги. Гага питается на морской литорали, где основную ее пищу составляют мелкие улитки из рода Littorina. В связи с особенностями жизненного цикла М. pygmaeus с каждым съеденным моллюском птица получает сотни метацеркарий, в кишечнике взрослой гаги насчитывается до 20— 30 тыс. паразитов. Даже у двухнедельных птенцов число сосальщиков определяется 2—3 тыс. В результате взрослые птицы болеют, а птенцы гибнут. На Белом море, где гаги обычны и охраняются, периодически возникают эпизоотии, сильно сокращающие численность этих птиц.
Дигенетические сосальщики, или трематоды (лат. Digenea) — класс паразитических плоских червей (Plathelminthes). Их жизненный цикл, за редкими исключениями, протекает в нескольких хозяевах и сопровождается закономерным чередованием не менее трёх поколений. Описано около 7200 видов. Некоторые трематоды (около 40 видов) — паразиты человека, вызывающие опасные заболевания — трематодозы, например, шистосомоз и описторхоз. К трематодам относятся печёночная двуустка (Fasciola hepatica), кошачья двуустка (Opisthorchis felineus), шистосома (Schistosoma), лейкохлоридий парадоксальный (Leucochloridium paradoxum).
В отличие от отечественной научной традиции, в современной англоязычной научной литературе название Trematoda используют для обозначения таксона, объединяющего аспидогастров (Aspidogastrea) и собственно дигенетических сосальщиков, за которыми сохраняется название Digenea. Это противоречие носит сугубо номенклатурный характер. / (Википедия)
Книги:
 Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987
Жизнь животных. Том 1. Простейшие. Кишечнополостные. Черви / В. Е. Соколов - 1987
 Трематоды животных и человека. Основы трематодологии. Том 1 (Общая трематодология; Atractotrematidae -- Echinostomatidae) / Скрябин К.И. - 1947
Трематоды животных и человека. Основы трематодологии. Том 1 (Общая трематодология; Atractotrematidae -- Echinostomatidae) / Скрябин К.И. - 1947
 Трематоды животных и человека. Основы трематодологии. Том 2 (Fasciolidae -- Lecithodendriidae) / Скрябин К.И. - 1948
Трематоды животных и человека. Основы трематодологии. Том 2 (Fasciolidae -- Lecithodendriidae) / Скрябин К.И. - 1948
 Трематоды животных и человека. Основы трематодологии. Том 3 (Paramphistomatata) / Скрябин К.И. - 1949
Трематоды животных и человека. Основы трематодологии. Том 3 (Paramphistomatata) / Скрябин К.И. - 1949
 Трематоды животных и человека. Основы трематодологии. Том 4 (Cephalogonimidae -- Cyclocoeliidae) / Скрябин К.И. - 1950
Трематоды животных и человека. Основы трематодологии. Том 4 (Cephalogonimidae -- Cyclocoeliidae) / Скрябин К.И. - 1950
 Трематоды животных и человека. Основы трематодологии. Том 5 (Schistosomatata) / Скрябин К.И. - 1951
Трематоды животных и человека. Основы трематодологии. Том 5 (Schistosomatata) / Скрябин К.И. - 1951
 Трематоды животных и человека. Основы трематодологии. Том 6 (Aspidogastrea, Heterophyoidea, Microphallidae) / Скрябин К.И. - 1952
Трематоды животных и человека. Основы трематодологии. Том 6 (Aspidogastrea, Heterophyoidea, Microphallidae) / Скрябин К.И. - 1952
 Трематоды животных и человека. Основы трематодологии. Том 7 (Cephaloporidae, Monodhelmidae, Dicrocoeliidae, Gorgoderidae) / Скрябин К.И. - 1952
Трематоды животных и человека. Основы трематодологии. Том 7 (Cephaloporidae, Monodhelmidae, Dicrocoeliidae, Gorgoderidae) / Скрябин К.И. - 1952
 Трематоды животных и человека. Основы трематодологии. Том 8 (Notocotylata, Gorgoderidae) / Скрябин К.И. - 1953
Трематоды животных и человека. Основы трематодологии. Том 8 (Notocotylata, Gorgoderidae) / Скрябин К.И. - 1953
 Трематоды животных и человека. Основы трематодологии. Том 9 (Opistolebetidae, Acanthocolpidae, Hemiuridae, Lecithasteridae, Dinuridae) / Скрябин К.И. - 1954
Трематоды животных и человека. Основы трематодологии. Том 9 (Opistolebetidae, Acanthocolpidae, Hemiuridae, Lecithasteridae, Dinuridae) / Скрябин К.И. - 1954
 Трематоды животных и человека. Основы трематодологии. Том 10 (Pronocephalidae, Megasolenidae, Acanthostomatidae, Gymnophallidae, Hemiurata) / Скрябин К.И. - 1955
Трематоды животных и человека. Основы трематодологии. Том 10 (Pronocephalidae, Megasolenidae, Acanthostomatidae, Gymnophallidae, Hemiurata) / Скрябин К.И. - 1955
 Трематоды животных и человека. Основы трематодологии. Том 11 (Didymozoata, Monorchidae, Halipegidae) / Скрябин К.И. - 1955
Трематоды животных и человека. Основы трематодологии. Том 11 (Didymozoata, Monorchidae, Halipegidae) / Скрябин К.И. - 1955
 Трематоды животных и человека. Основы трематодологии. Том 12 (Haploporidae, Echinostomatidae) / Скрябин К.И. - 1956
Трематоды животных и человека. Основы трематодологии. Том 12 (Haploporidae, Echinostomatidae) / Скрябин К.И. - 1956
 Трематоды животных и человека. Основы трематодологии. Том 13 (Zoogonidae, Steganodermatidae, Fellodistomatidae) / Скрябин К.И. - 1957
Трематоды животных и человека. Основы трематодологии. Том 13 (Zoogonidae, Steganodermatidae, Fellodistomatidae) / Скрябин К.И. - 1957
 Трематоды животных и человека. Основы трематодологии. Том 14 (Plagiorchioidea, Azygiata) / Скрябин К.И. - 1958
Трематоды животных и человека. Основы трематодологии. Том 14 (Plagiorchioidea, Azygiata) / Скрябин К.И. - 1958
 Трематоды животных и человека. Основы трематодологии. Том 15 (Thapariellidae, Trematobrienidae, Transversotrematidae, Prosogonotrematidae, Maseniidae, Deropristidae, Opecoelidae) / Скрябин К.И. - 1958
Трематоды животных и человека. Основы трематодологии. Том 15 (Thapariellidae, Trematobrienidae, Transversotrematidae, Prosogonotrematidae, Maseniidae, Deropristidae, Opecoelidae) / Скрябин К.И. - 1958
 Трематоды животных и человека. Основы трематодологии. Том 16 (Faustulida, Schistorchidae, Callodistomatidae) / Скрябин К.И. - 1959
Трематоды животных и человека. Основы трематодологии. Том 16 (Faustulida, Schistorchidae, Callodistomatidae) / Скрябин К.И. - 1959
 Трематоды животных и человека. Основы трематодологии. Том 17 (Hemiurata (окончание)) / Скрябин К.И. - 1960
Трематоды животных и человека. Основы трематодологии. Том 17 (Hemiurata (окончание)) / Скрябин К.И. - 1960
 Трематоды животных и человека. Основы трематодологии. Том 18 (Lepocriadioidea, Strigeata, Plagiorchioidea) / Скрябин К.И. - 1960
Трематоды животных и человека. Основы трематодологии. Том 18 (Lepocriadioidea, Strigeata, Plagiorchioidea) / Скрябин К.И. - 1960
 Трематоды животных и человека. Основы трематодологии. Том 19 (Prosthogonitidae, Plagiorchidae, Strigeidida) / Скрябин К.И. - 1961
Трематоды животных и человека. Основы трематодологии. Том 19 (Prosthogonitidae, Plagiorchidae, Strigeidida) / Скрябин К.И. - 1961
 Трематоды животных и человека. Основы трематодологии. Том 20 (Bucephalata) / Скрябин К.И. - 1962
Трематоды животных и человека. Основы трематодологии. Том 20 (Bucephalata) / Скрябин К.И. - 1962
 Трематоды животных и человека. Основы трематодологии. Том 21 (Brachycoeliidae, Pachypsolidae, Mesometridae, Orientocreadiidae, Telorchidae, Microphallidae) / Скрябин К.И. - 1963
Трематоды животных и человека. Основы трематодологии. Том 21 (Brachycoeliidae, Pachypsolidae, Mesometridae, Orientocreadiidae, Telorchidae, Microphallidae) / Скрябин К.И. - 1963
 Трематоды животных и человека. Основы трематодологии. Том 22 (Tandanicolidae, Ornithotrematidae, Parorchidae, Eumegacetidae, Allocreadioidea) / Скрябин К.И. - 1966
Трематоды животных и человека. Основы трематодологии. Том 22 (Tandanicolidae, Ornithotrematidae, Parorchidae, Eumegacetidae, Allocreadioidea) / Скрябин К.И. - 1966
 Трематоды животных и человека. Основы трематодологии. Том 23 (Dicrocoeliidae, Renicolidae, Pleurogenidae) / Скрябин К.И. - 1970
Трематоды животных и человека. Основы трематодологии. Том 23 (Dicrocoeliidae, Renicolidae, Pleurogenidae) / Скрябин К.И. - 1970
 Трематоды животных и человека. Основы трематодологии. Том 24 (Plagiorchidae, Strigeidida, Sphaerostomatinae, Stomylotrematidae, Ochetosomatidae) / Скрябин К.И. - 1971
Трематоды животных и человека. Основы трематодологии. Том 24 (Plagiorchidae, Strigeidida, Sphaerostomatinae, Stomylotrematidae, Ochetosomatidae) / Скрябин К.И. - 1971
 Трематоды животных и человека. Основы трематодологии. Том 25 (Wallinidae, Strigeidida, Allassogonoporidae, Cryptoporidae, указатель) / Скрябин К.И. - 1974
Трематоды животных и человека. Основы трематодологии. Том 25 (Wallinidae, Strigeidida, Allassogonoporidae, Cryptoporidae, указатель) / Скрябин К.И. - 1974
 Трематоды животных и человека. Основы трематодологии. Том 26 (Paragonimidae; Указатель видов трематод) / Скрябин К.И. - 1978
Трематоды животных и человека. Основы трематодологии. Том 26 (Paragonimidae; Указатель видов трематод) / Скрябин К.И. - 1978



 Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
Зоология беспозвоночных. Функциональные и эволюционные аспекты. В 4 томах. / Рупперт Э.Э., Фокс Р.С., Барнс Р.Д. - 2008
 Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423
Вестхайде В., Ригер Р. Зоология беспозвоночных. = Spezielle Zoology. Teil 1: Einzeller und Wirbellose Tiere / пер. с нем. О. Н. Бёллинг, С. М. Ляпкова, А. В. Михеев, О. Г. Манылов, А. А. Оскольский, А. В. Филиппова, А. В. Чесунов; под ред. А. В. Чесунова. — М.: Товарищество научных изданий КМК, 2008. — С. 366—423
 Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989
Биологический энциклопедический словарь под редакцией М. С. Гилярова и др., М., изд. Советская Энциклопедия, 1989